Research ArticleImmunologyInfectious disease
Open Access | ![]() 10.1172/jci.insight.189134
10.1172/jci.insight.189134
Bcl6 expression is associated with a distinct immune landscape and spatial transcriptome in COVID-19
Cloé Brenna,1 Bernat Bramon Mora,2 Kalliopi Ioannidou,1 Julien Bodelet,3,4 Mia L. Siebmanns,1 Simon Burgermeister,1 Spiros Georgakis,1 Michail Orfanakis,1 Yannick D. Muller,5 Nazanin Sédille,6 Matthew J. Feinstein,7 Jon W. Lomasney,7 Oliver Y. Chén,3,4 Giuseppe Pantaleo,5 Sabina Berezowska,1 Laurence de Leval,1 Raphael Gottardo,2 and Constantinos Petrovas1
1Institute of Pathology, Department of Laboratory Medicine and Pathology, and
2Biomedical Data Science Center, Lausanne University Hospital (CHUV), Lausanne, Switzerland.
3Bioinformatics Platform, Department of Laboratory Medicine and Pathology, CHUV, Lausanne, Switzerland.
4Faculty of Biology and Medicine, University of Lausanne, Lausanne, Switzerland.
5Service of Immunology and Allergy, Department of Medicine, and
6Service of Clinical Chemistry, Department of Laboratory Medicine and Pathology, CHUV, Lausanne, Switzerland.
7Feinberg School of Medicine of Northwestern University, Chicago, Illinois, USA.
Address correspondence to: Constantinos Petrovas, Institute of Pathology, Department of Laboratory Medicine and Pathology, Lausanne University Hospital and Lausanne University, Rue du Bugnon 25, CH - 1011, Lausanne, Switzerland. Email: Konstantinos.Petrovas@chuv.ch.
Find articles by Brenna, C. in: PubMed | Google Scholar
1Institute of Pathology, Department of Laboratory Medicine and Pathology, and
2Biomedical Data Science Center, Lausanne University Hospital (CHUV), Lausanne, Switzerland.
3Bioinformatics Platform, Department of Laboratory Medicine and Pathology, CHUV, Lausanne, Switzerland.
4Faculty of Biology and Medicine, University of Lausanne, Lausanne, Switzerland.
5Service of Immunology and Allergy, Department of Medicine, and
6Service of Clinical Chemistry, Department of Laboratory Medicine and Pathology, CHUV, Lausanne, Switzerland.
7Feinberg School of Medicine of Northwestern University, Chicago, Illinois, USA.
Address correspondence to: Constantinos Petrovas, Institute of Pathology, Department of Laboratory Medicine and Pathology, Lausanne University Hospital and Lausanne University, Rue du Bugnon 25, CH - 1011, Lausanne, Switzerland. Email: Konstantinos.Petrovas@chuv.ch.
Find articles by Bramon Mora, B. in: PubMed | Google Scholar
1Institute of Pathology, Department of Laboratory Medicine and Pathology, and
2Biomedical Data Science Center, Lausanne University Hospital (CHUV), Lausanne, Switzerland.
3Bioinformatics Platform, Department of Laboratory Medicine and Pathology, CHUV, Lausanne, Switzerland.
4Faculty of Biology and Medicine, University of Lausanne, Lausanne, Switzerland.
5Service of Immunology and Allergy, Department of Medicine, and
6Service of Clinical Chemistry, Department of Laboratory Medicine and Pathology, CHUV, Lausanne, Switzerland.
7Feinberg School of Medicine of Northwestern University, Chicago, Illinois, USA.
Address correspondence to: Constantinos Petrovas, Institute of Pathology, Department of Laboratory Medicine and Pathology, Lausanne University Hospital and Lausanne University, Rue du Bugnon 25, CH - 1011, Lausanne, Switzerland. Email: Konstantinos.Petrovas@chuv.ch.
Find articles by Ioannidou, K. in: PubMed | Google Scholar
1Institute of Pathology, Department of Laboratory Medicine and Pathology, and
2Biomedical Data Science Center, Lausanne University Hospital (CHUV), Lausanne, Switzerland.
3Bioinformatics Platform, Department of Laboratory Medicine and Pathology, CHUV, Lausanne, Switzerland.
4Faculty of Biology and Medicine, University of Lausanne, Lausanne, Switzerland.
5Service of Immunology and Allergy, Department of Medicine, and
6Service of Clinical Chemistry, Department of Laboratory Medicine and Pathology, CHUV, Lausanne, Switzerland.
7Feinberg School of Medicine of Northwestern University, Chicago, Illinois, USA.
Address correspondence to: Constantinos Petrovas, Institute of Pathology, Department of Laboratory Medicine and Pathology, Lausanne University Hospital and Lausanne University, Rue du Bugnon 25, CH - 1011, Lausanne, Switzerland. Email: Konstantinos.Petrovas@chuv.ch.
Find articles by Bodelet, J. in: PubMed | Google Scholar
1Institute of Pathology, Department of Laboratory Medicine and Pathology, and
2Biomedical Data Science Center, Lausanne University Hospital (CHUV), Lausanne, Switzerland.
3Bioinformatics Platform, Department of Laboratory Medicine and Pathology, CHUV, Lausanne, Switzerland.
4Faculty of Biology and Medicine, University of Lausanne, Lausanne, Switzerland.
5Service of Immunology and Allergy, Department of Medicine, and
6Service of Clinical Chemistry, Department of Laboratory Medicine and Pathology, CHUV, Lausanne, Switzerland.
7Feinberg School of Medicine of Northwestern University, Chicago, Illinois, USA.
Address correspondence to: Constantinos Petrovas, Institute of Pathology, Department of Laboratory Medicine and Pathology, Lausanne University Hospital and Lausanne University, Rue du Bugnon 25, CH - 1011, Lausanne, Switzerland. Email: Konstantinos.Petrovas@chuv.ch.
Find articles by Siebmanns, M. in: PubMed | Google Scholar
1Institute of Pathology, Department of Laboratory Medicine and Pathology, and
2Biomedical Data Science Center, Lausanne University Hospital (CHUV), Lausanne, Switzerland.
3Bioinformatics Platform, Department of Laboratory Medicine and Pathology, CHUV, Lausanne, Switzerland.
4Faculty of Biology and Medicine, University of Lausanne, Lausanne, Switzerland.
5Service of Immunology and Allergy, Department of Medicine, and
6Service of Clinical Chemistry, Department of Laboratory Medicine and Pathology, CHUV, Lausanne, Switzerland.
7Feinberg School of Medicine of Northwestern University, Chicago, Illinois, USA.
Address correspondence to: Constantinos Petrovas, Institute of Pathology, Department of Laboratory Medicine and Pathology, Lausanne University Hospital and Lausanne University, Rue du Bugnon 25, CH - 1011, Lausanne, Switzerland. Email: Konstantinos.Petrovas@chuv.ch.
Find articles by Burgermeister, S. in: PubMed | Google Scholar
1Institute of Pathology, Department of Laboratory Medicine and Pathology, and
2Biomedical Data Science Center, Lausanne University Hospital (CHUV), Lausanne, Switzerland.
3Bioinformatics Platform, Department of Laboratory Medicine and Pathology, CHUV, Lausanne, Switzerland.
4Faculty of Biology and Medicine, University of Lausanne, Lausanne, Switzerland.
5Service of Immunology and Allergy, Department of Medicine, and
6Service of Clinical Chemistry, Department of Laboratory Medicine and Pathology, CHUV, Lausanne, Switzerland.
7Feinberg School of Medicine of Northwestern University, Chicago, Illinois, USA.
Address correspondence to: Constantinos Petrovas, Institute of Pathology, Department of Laboratory Medicine and Pathology, Lausanne University Hospital and Lausanne University, Rue du Bugnon 25, CH - 1011, Lausanne, Switzerland. Email: Konstantinos.Petrovas@chuv.ch.
Find articles by Georgakis, S. in: PubMed | Google Scholar
1Institute of Pathology, Department of Laboratory Medicine and Pathology, and
2Biomedical Data Science Center, Lausanne University Hospital (CHUV), Lausanne, Switzerland.
3Bioinformatics Platform, Department of Laboratory Medicine and Pathology, CHUV, Lausanne, Switzerland.
4Faculty of Biology and Medicine, University of Lausanne, Lausanne, Switzerland.
5Service of Immunology and Allergy, Department of Medicine, and
6Service of Clinical Chemistry, Department of Laboratory Medicine and Pathology, CHUV, Lausanne, Switzerland.
7Feinberg School of Medicine of Northwestern University, Chicago, Illinois, USA.
Address correspondence to: Constantinos Petrovas, Institute of Pathology, Department of Laboratory Medicine and Pathology, Lausanne University Hospital and Lausanne University, Rue du Bugnon 25, CH - 1011, Lausanne, Switzerland. Email: Konstantinos.Petrovas@chuv.ch.
Find articles by Orfanakis, M. in: PubMed | Google Scholar
1Institute of Pathology, Department of Laboratory Medicine and Pathology, and
2Biomedical Data Science Center, Lausanne University Hospital (CHUV), Lausanne, Switzerland.
3Bioinformatics Platform, Department of Laboratory Medicine and Pathology, CHUV, Lausanne, Switzerland.
4Faculty of Biology and Medicine, University of Lausanne, Lausanne, Switzerland.
5Service of Immunology and Allergy, Department of Medicine, and
6Service of Clinical Chemistry, Department of Laboratory Medicine and Pathology, CHUV, Lausanne, Switzerland.
7Feinberg School of Medicine of Northwestern University, Chicago, Illinois, USA.
Address correspondence to: Constantinos Petrovas, Institute of Pathology, Department of Laboratory Medicine and Pathology, Lausanne University Hospital and Lausanne University, Rue du Bugnon 25, CH - 1011, Lausanne, Switzerland. Email: Konstantinos.Petrovas@chuv.ch.
Find articles by Muller, Y. in: PubMed | Google Scholar
1Institute of Pathology, Department of Laboratory Medicine and Pathology, and
2Biomedical Data Science Center, Lausanne University Hospital (CHUV), Lausanne, Switzerland.
3Bioinformatics Platform, Department of Laboratory Medicine and Pathology, CHUV, Lausanne, Switzerland.
4Faculty of Biology and Medicine, University of Lausanne, Lausanne, Switzerland.
5Service of Immunology and Allergy, Department of Medicine, and
6Service of Clinical Chemistry, Department of Laboratory Medicine and Pathology, CHUV, Lausanne, Switzerland.
7Feinberg School of Medicine of Northwestern University, Chicago, Illinois, USA.
Address correspondence to: Constantinos Petrovas, Institute of Pathology, Department of Laboratory Medicine and Pathology, Lausanne University Hospital and Lausanne University, Rue du Bugnon 25, CH - 1011, Lausanne, Switzerland. Email: Konstantinos.Petrovas@chuv.ch.
Find articles by Sédille, N. in: PubMed | Google Scholar
1Institute of Pathology, Department of Laboratory Medicine and Pathology, and
2Biomedical Data Science Center, Lausanne University Hospital (CHUV), Lausanne, Switzerland.
3Bioinformatics Platform, Department of Laboratory Medicine and Pathology, CHUV, Lausanne, Switzerland.
4Faculty of Biology and Medicine, University of Lausanne, Lausanne, Switzerland.
5Service of Immunology and Allergy, Department of Medicine, and
6Service of Clinical Chemistry, Department of Laboratory Medicine and Pathology, CHUV, Lausanne, Switzerland.
7Feinberg School of Medicine of Northwestern University, Chicago, Illinois, USA.
Address correspondence to: Constantinos Petrovas, Institute of Pathology, Department of Laboratory Medicine and Pathology, Lausanne University Hospital and Lausanne University, Rue du Bugnon 25, CH - 1011, Lausanne, Switzerland. Email: Konstantinos.Petrovas@chuv.ch.
Find articles by
Feinstein, M.
in:
PubMed
|
Google Scholar
|

1Institute of Pathology, Department of Laboratory Medicine and Pathology, and
2Biomedical Data Science Center, Lausanne University Hospital (CHUV), Lausanne, Switzerland.
3Bioinformatics Platform, Department of Laboratory Medicine and Pathology, CHUV, Lausanne, Switzerland.
4Faculty of Biology and Medicine, University of Lausanne, Lausanne, Switzerland.
5Service of Immunology and Allergy, Department of Medicine, and
6Service of Clinical Chemistry, Department of Laboratory Medicine and Pathology, CHUV, Lausanne, Switzerland.
7Feinberg School of Medicine of Northwestern University, Chicago, Illinois, USA.
Address correspondence to: Constantinos Petrovas, Institute of Pathology, Department of Laboratory Medicine and Pathology, Lausanne University Hospital and Lausanne University, Rue du Bugnon 25, CH - 1011, Lausanne, Switzerland. Email: Konstantinos.Petrovas@chuv.ch.
Find articles by
Lomasney, J.
in:
PubMed
|
Google Scholar
|
1Institute of Pathology, Department of Laboratory Medicine and Pathology, and
2Biomedical Data Science Center, Lausanne University Hospital (CHUV), Lausanne, Switzerland.
3Bioinformatics Platform, Department of Laboratory Medicine and Pathology, CHUV, Lausanne, Switzerland.
4Faculty of Biology and Medicine, University of Lausanne, Lausanne, Switzerland.
5Service of Immunology and Allergy, Department of Medicine, and
6Service of Clinical Chemistry, Department of Laboratory Medicine and Pathology, CHUV, Lausanne, Switzerland.
7Feinberg School of Medicine of Northwestern University, Chicago, Illinois, USA.
Address correspondence to: Constantinos Petrovas, Institute of Pathology, Department of Laboratory Medicine and Pathology, Lausanne University Hospital and Lausanne University, Rue du Bugnon 25, CH - 1011, Lausanne, Switzerland. Email: Konstantinos.Petrovas@chuv.ch.
Find articles by Chén, O. in: PubMed | Google Scholar
1Institute of Pathology, Department of Laboratory Medicine and Pathology, and
2Biomedical Data Science Center, Lausanne University Hospital (CHUV), Lausanne, Switzerland.
3Bioinformatics Platform, Department of Laboratory Medicine and Pathology, CHUV, Lausanne, Switzerland.
4Faculty of Biology and Medicine, University of Lausanne, Lausanne, Switzerland.
5Service of Immunology and Allergy, Department of Medicine, and
6Service of Clinical Chemistry, Department of Laboratory Medicine and Pathology, CHUV, Lausanne, Switzerland.
7Feinberg School of Medicine of Northwestern University, Chicago, Illinois, USA.
Address correspondence to: Constantinos Petrovas, Institute of Pathology, Department of Laboratory Medicine and Pathology, Lausanne University Hospital and Lausanne University, Rue du Bugnon 25, CH - 1011, Lausanne, Switzerland. Email: Konstantinos.Petrovas@chuv.ch.
Find articles by Pantaleo, G. in: PubMed | Google Scholar
1Institute of Pathology, Department of Laboratory Medicine and Pathology, and
2Biomedical Data Science Center, Lausanne University Hospital (CHUV), Lausanne, Switzerland.
3Bioinformatics Platform, Department of Laboratory Medicine and Pathology, CHUV, Lausanne, Switzerland.
4Faculty of Biology and Medicine, University of Lausanne, Lausanne, Switzerland.
5Service of Immunology and Allergy, Department of Medicine, and
6Service of Clinical Chemistry, Department of Laboratory Medicine and Pathology, CHUV, Lausanne, Switzerland.
7Feinberg School of Medicine of Northwestern University, Chicago, Illinois, USA.
Address correspondence to: Constantinos Petrovas, Institute of Pathology, Department of Laboratory Medicine and Pathology, Lausanne University Hospital and Lausanne University, Rue du Bugnon 25, CH - 1011, Lausanne, Switzerland. Email: Konstantinos.Petrovas@chuv.ch.
Find articles by Berezowska, S. in: PubMed | Google Scholar
1Institute of Pathology, Department of Laboratory Medicine and Pathology, and
2Biomedical Data Science Center, Lausanne University Hospital (CHUV), Lausanne, Switzerland.
3Bioinformatics Platform, Department of Laboratory Medicine and Pathology, CHUV, Lausanne, Switzerland.
4Faculty of Biology and Medicine, University of Lausanne, Lausanne, Switzerland.
5Service of Immunology and Allergy, Department of Medicine, and
6Service of Clinical Chemistry, Department of Laboratory Medicine and Pathology, CHUV, Lausanne, Switzerland.
7Feinberg School of Medicine of Northwestern University, Chicago, Illinois, USA.
Address correspondence to: Constantinos Petrovas, Institute of Pathology, Department of Laboratory Medicine and Pathology, Lausanne University Hospital and Lausanne University, Rue du Bugnon 25, CH - 1011, Lausanne, Switzerland. Email: Konstantinos.Petrovas@chuv.ch.
Find articles by
de Leval, L.
in:
PubMed
|
Google Scholar
|
1Institute of Pathology, Department of Laboratory Medicine and Pathology, and
2Biomedical Data Science Center, Lausanne University Hospital (CHUV), Lausanne, Switzerland.
3Bioinformatics Platform, Department of Laboratory Medicine and Pathology, CHUV, Lausanne, Switzerland.
4Faculty of Biology and Medicine, University of Lausanne, Lausanne, Switzerland.
5Service of Immunology and Allergy, Department of Medicine, and
6Service of Clinical Chemistry, Department of Laboratory Medicine and Pathology, CHUV, Lausanne, Switzerland.
7Feinberg School of Medicine of Northwestern University, Chicago, Illinois, USA.
Address correspondence to: Constantinos Petrovas, Institute of Pathology, Department of Laboratory Medicine and Pathology, Lausanne University Hospital and Lausanne University, Rue du Bugnon 25, CH - 1011, Lausanne, Switzerland. Email: Konstantinos.Petrovas@chuv.ch.
Find articles by Gottardo, R. in: PubMed | Google Scholar
1Institute of Pathology, Department of Laboratory Medicine and Pathology, and
2Biomedical Data Science Center, Lausanne University Hospital (CHUV), Lausanne, Switzerland.
3Bioinformatics Platform, Department of Laboratory Medicine and Pathology, CHUV, Lausanne, Switzerland.
4Faculty of Biology and Medicine, University of Lausanne, Lausanne, Switzerland.
5Service of Immunology and Allergy, Department of Medicine, and
6Service of Clinical Chemistry, Department of Laboratory Medicine and Pathology, CHUV, Lausanne, Switzerland.
7Feinberg School of Medicine of Northwestern University, Chicago, Illinois, USA.
Address correspondence to: Constantinos Petrovas, Institute of Pathology, Department of Laboratory Medicine and Pathology, Lausanne University Hospital and Lausanne University, Rue du Bugnon 25, CH - 1011, Lausanne, Switzerland. Email: Konstantinos.Petrovas@chuv.ch.
Find articles by Petrovas, C. in: PubMed | Google Scholar
Published September 9, 2025 - More info
JCI Insight. 2025;10(20):e189134. https://doi.org/10.1172/jci.insight.189134.
© 2025 Cloé Brenna et al. This work is licensed under the Creative Commons Attribution 4.0 International License. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
Received: November 11, 2024; Accepted: September 4, 2025

-
Results
COVID-19 infection induces heterogeneous RF profiling in LD-LNs. We started our investigation by analyzing F/GC B cell subsets in LD-LNs from COVID-19–infected individuals from 2 centers (CHUV and Northwestern University) (Table 1 and Supplemental Table 1) by applying a mIF (Supplemental Tables 2 and 3) that allows for the simultaneous detection of CD20, Bcl6, and Ki67 (Figure 1 and Supplemental Figure 1). HLN-Controls from non-COVID-19–infected donors (n = 4) were also included in the analysis (Table 1 and Supplemental Figure 2).
 Figure 1
Figure 1COVID-19 infection induces diverse and heterogeneous reactive follicles in LD-LNs. (A) Representative multiplex fluorescence images showing follicular areas from RF-Bcl6no/lo (Donor 2), RF-Bcl6int (Donor 16), and RF-Bcl6hi (Donor 17) tissues with CD20hi (blue), Ki67hi (magenta), Bcl6hi (green), and merged images, highlighting differences in follicular reactivity. Scale bars: 100 μm. (B) Histocytometry gating scheme for B cell subset identification based on CD20hi/dim density, with prevalence calculated for all follicles combined (All F). (C) Dot plots displaying absolute counts of B cell subsets (CD20hi/dim, CD20hi/dimKi67hi, CD20hi/dimKi67hiBcl6hi) in individual follicles from representative donors — RF-Bcl6no/lo as Donor 4, RF-Bcl6int as Donor 13, RF-Bcl6hi as Donor 22 — illustrating variations in B cell proliferation. The fourth panel summarizes total follicular area CD20hi/dimKi67hiBcl6hi cell counts across the 4 groups (RF-Bcl6no/lo, purple; RF-Bcl6int, pink; RF-Bcl6hi, green), with blue diamonds representing HLN-Controls from non-COVID-19–infected donors. (D) Dot plot displaying normalized follicular areas across the 3 RF groups: RF-Bcl6no/lo, purple; RF-Bcl6int, pink; and RF-Bcl6hi, green. Control HLNs from non-COVID-19–infected donors are represented by blue diamonds, highlighting differences in follicular size and maturity. (E) Dot plots showing the B cell subset densities (cells/μm²) — CD20hi/dim, CD20hi/dim Ki67hi, and CD20hi/dimKi67hiBcl6hi — measured in merged follicular areas from each donor. Groups are color-coded as follows: RF-Bcl6no/lo, purple; RF-Bcl6int, pink; RF-Bcl6hi, green; and HLNs from non-COVID-19–infected controls, blue. Light blue dot within the RF-Bcl6no/lo group corresponds to an axillary LN (Donor 25); yellow dot corresponds to para-aortic and mediastinal LNs (Donor 29); and gray dots within the RF-Bcl6int group represent mediastinal LN (Donors 24-a and 24-b). Statistical comparisons were performed using the Mann-Whitney U test; *P ≤ 0.05, **P ≤ 0.01, ***P ≤ 0.001, ****P ≤ 0.0001. Raw P values are shown on the graph, and both raw and FDR-adjusted P values are reported in Supplemental Table 11.
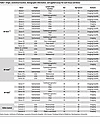 Table 1
Table 1Origin, anatomical location, demographic information, and applied assays for each tissue and donor
Follicular areas, identified by the density of CD20hi/dim cells, were further analyzed for B cell subsets using histocytometry analysis for all combined follicles (total follicular area [All F]) (Figure 1B). Visual inspection of raw mIF images, along with quantitative analysis of Bcl6hi B cell counts, either per follicular area or total follicular area per tissue, revealed a high heterogeneity of follicular Bcl6 expression within individual LNs, with mixed levels observed among follicles (Figure 1C; Supplemental Figure 3, A–C; and Supplemental Figure 4A).
Based on the Bcl6 profiling, 3 distinct tissue groups were identified: RF-Bcl6no/lo (mean ± SD: 48.3 ± 23.9 follicles analyzed), RF-Bcl6int (50.8 ± 24.9 follicles analyzed), and RF-Bcl6hi (66.3 ± 25.2 follicles analyzed) (Figure 1C; Supplemental Figure 3, A–C; and Supplemental Figure 4A). No significant difference in the number of follicles analyzed among the 3 groups was observed, as assessed by a Mann-Whitney U test (Supplemental Table 4).
The time interval between death and autopsy procedure did not differ significantly between groups (CHUV samples), with an average of 19.86 ± 12.26 hours (RF-Bcl6no/lo), 35.53 ± 24.70 hours (RF-Bcl6int), and 33.22 ± 26.72 hours (RF-Bcl6hi) (Supplemental Table 5). A Mann-Whitney U test showed no significant differences between death and autopsy procedure between RF-Bcl6no/lo and RF-Bcl6int (P = 0.1451), RF-Bcl6no/lo and RF-Bcl6hi (P = 0.3283), or RF-Bcl6int and RF-Bcl6hi (P > 0.9999), confirming no influence on RF-Bcl6 expression patterns. Furthermore, no consistent association was observed between immunosuppressive treatments and Bcl6 expression levels across donors, suggesting that local immune dynamics, rather than systemic immunosuppression alone, may drive the heterogeneity in RF-Bcl6 groups (Table 2). Notably, the RF-Bcl6no/lo group displayed Ki67 expression, indicating active proliferation and confirming these as RF rather than primary or resting follicles.
Analysis of RF morphological features revealed that individual RFs exhibited a significantly larger area in both RF-Bcl6int and RF-Bcl6hi groups when compared with RF-Bcl6no/lo and HLN-Controls from non-COVID-19–infected donors (Figure 1D). Although an increase in follicular size can be observed in various reactive contexts, our data indicate that this enlargement is particularly associated with higher Bcl6 expression in the setting of COVID-19, potentially reflecting follicular expansion and/or enhanced maturation of GC responses in the former 2 groups. No significant differences were observed in other morphological features analyzed.
Next, the cell densities (normalized cell counts per μm²) of specific B cell subsets were calculated for all available tissues. Despite similar cell densities of bulk CD20hi/dim B cells across the HLN-Controls and the 3 groups, RF-Bcl6int and especially RF-Bcl6hi tissues were characterized by significantly higher densities of proliferating Ki67hi and CD20hi/dimKi67hiBcl6hi B cells (Figure 1E). A similar profile was observed when the frequency of B cell subsets was calculated (Supplemental Figure 4A).
Then, the pattern of the follicular DC (FDC) network, as an additional surrogate of mature RFs, was evaluated (Supplemental Figure 4B). We found a diverse expression of FDC in most RFs from both RF-Bcl6no/lo and RF-Bcl6hi tissues. However, the FDC staining pattern varied between the 2 groups, with a connected network pattern more frequently observed in RF-Bcl6hi tissues, while unconnected FDC+ events, a sign of less active and mature RFs, were commonly seen in RF-Bcl6no/lo tissues (Supplemental Figure 4B). Altogether, our data suggest that RFs, as indicated by the cell densities of Ki67hi and Bcl6hi B cells, in LD-LNs from COVID-19–infected individuals are highly diverse and heterogeneous.
We next assessed the relationship between RF-Bcl6 expression and serological markers taken during hospitalization (CHUV donors) using a mixed linear regression model to account for repeated measures and donor variability (Supplemental Table 6). A trend toward higher CRP levels was observed in RF-Bcl6no/lo compared with RF-Bcl6hi (β = –61.56, P = 0.257), though leukocyte and lymphocyte counts did not differ significantly (Supplemental Table 6). We then examined whether RF-Bcl6 expression varied in relation to the estimated time after infection, as estimated by the duration of hospitalization. Given the cross-sectional nature of the study, this analysis does not infer longitudinal changes within individuals but assesses whether longer hospitalization is associated with differences in Bcl6 expression. No significant changes in RF-Bcl6 expression were observed over time in RF-Bcl6no/lo (P = 0.927) or RF-Bcl6hi (P = 0.903) groups (Supplemental Figure 4C and Supplemental Table 7), suggesting that Bcl6 expression does not progressively increase during hospitalization. However, a significant decline was observed in the RF-Bcl6int group (β = –60.175, P = 0.013) (Supplemental Table 7), while at the general level only a modest, nonsignificant decrease was detected (β = –0.00446, P = 0.179) (Supplemental Table 7). Overall, our data suggest that the heterogeneity of LD-LNs RF-Bcl6 expression likely reflects the intrinsic capacity of the immune system to mount mature GC-RFs rather than a direct consequence of hospitalization duration or post-infection timing.
RF-Bcl6hi LD-LNs harbor higher cell densities of proliferating CD57hi Tfh cell. Given the essential role of Tfh cells in the formation and maintenance of GC-RFs (5), we aimed to analyze their in situ cell densities across the 3 aforementioned groups. Different Tfh cell subsets were identified based on the expression of PD1, Bcl6, Ki67, and CD57 expression, a marker that defines a distinct Tfh cells subset (9, 10) (Figure 2 and Supplemental Figure 4D). We noticed an inconsistency in the coexpression of CD57 and CD4 in RFs among individuals, possibly reflecting a downregulation of the CD4 receptor, similar to the pattern observed for CD3 expression (9) in highly differentiated Tfh cells. Given this characteristic, we refined our gating strategy to avoid bias by identifying Tfh cells as PD1hi and PD1hiCD57hi (Figure 2B). To set relevant cut-off values, we used PD1 and CD57 expression levels in extrafollicular (EF) CD4 T cells as a reference and applied these thresholds across the entire F region (Figure 2B).
 Figure 2
Figure 2RF-Bcl6hi LD-LNs harbor higher densities of proliferating Bcl6hi and CD57hi Tfh cells. (A) Representative fluorescence images of follicular areas from RF-Bcl6no/lo (Donor 2), RF-Bcl6int (Donor 16), and RF-Bcl6hi (Donor 17) tissues showing PD1hi (orange), CD57hi (yellow), Bcl6hi (cyan), Ki67hi (white), and merged images, highlighting Tfh cell subset differences. Scale bars: 100 μm. (B) Histocytometry gating scheme for CD4 T cell subset identification within CD20hi/dim follicular areas, with prevalence calculated for all follicles combined (All F). (C) Dot plots showing absolute counts of Tfh cell subsets (PD1hi, PD1hiCD57hi, PD1hiKi67hi, PD1hiKi67hiBcl6hi) per follicle from representative donors (RF-Bcl6no/lo, Donor 4; RF-Bcl6int, Donor 13; RF-Bcl6hi, Donor 22). (D) Dot plots of normalized Tfh cell subset densities (cells/μm²) — PD1hi, PD1hi Ki67hi, PD1hiCD57hiBcl6hi, and PD1hiKi67hiBcl6hi — measured in merged follicular areas from each donor. Groups are color-coded as follows: RF-Bcl6no/lo (purple), RF-Bcl6int (pink), RF-Bcl6hi (green), and HLNs from non-COVID-19–infected controls (blue). Light blue dot within the RF-Bcl6no/lo group corresponds to an axillary LN (Donor 25); yellow dot corresponds to para-aortic and mediastinal LNs (Donor 29); gray dots within the RF-Bcl6int group represent mediastinal LN (Donors 24-a and 24-b). (E) SQL clustering projection of B cell and Tfh cell subsets based on the expression of CD20hi/dim, CD20hi/dimKi67hi, CD20hi/dimKi67hiBcl6hi, CD4hi, PD1hi, PD1hiKi67hi, PD1hiCD57hi, and PD1hiKi67hiBcl6hi. Data points are color-coded by RF group: RF-Bcl6no/lo (purple), RF-Bcl6int (pink), and RF-Bcl6hi (green). HLNs from non-COVID-19–infected control donors are shown as blue diamonds. The projection displays SQL Factor 1 versus SQL Factor 3, which capture the main variance across immune cell phenotypes and best separate the groups. Mann-Whitney U test; *P ≤ 0.05, **P ≤ 0.01, ***P ≤ 0.001, ****P ≤ 0.0001. Raw P values are shown on the graph, and both raw and FDR-adjusted P values are reported in Supplemental Table 11.
The calculation of absolute numbers of Bcl6+ Tfh cells across individual RFs revealed a distribution pattern similar to that observed for B cells (Figure 2C; Supplemental Figure 3, A–C; and Supplemental Figure 4A). A significantly higher cell density of PD1hi, PD1hiKi67hi, PD1hiCD57hi, and PD1hiKi67hiBcl6hi cells in RF-Bcl6int and RF-Bcl6hi compared with RF-Bcl6no/lo tissues was observed (Figure 2D), suggesting an association between Tfh cells differentiation and Bcl6-expression in these RFs. Unlike B cells, a significant difference between HLN-Controls from non-COVID-19–infected and RF-Bcl6hi LNs was found only for the PD1hiKi67hiBcl6hi cells (Figure 2D).
A similar trend was observed when analyzing the frequency of Tfh cell subsets (Supplemental Figure 4E). To further explore the relationship between Bcl6 expression and follicular immune cell composition, we applied the statistical quantile learning (SQL) method (11) for clustering analysis based on B cells and Tfh cells measurements. The SQL projection of adaptive immunity (Figure 2E) revealed a gradual transition in immune cell clustering that aligns with increasing Bcl6 expression.
Altogether, our data indicate that the expression of Bcl6 in LD-LNs from COVID-19 autopsies reflects a spectrum of follicular immune reactivity.
The increasing prevalence of Bcl6 is associated with an altered spatial distribution of RF immune cell subsets. Next, we analyzed the spatial distribution of key immune cell subsets within RFs in the RF-Bcl6int and RF-Bcl6hi groups by applying linear regression models and relevant algorithms.
A significant positive correlation was found between PD1hiCD57hi Tfh cells and CD20hi/dimKi67hiBcl6hi B cells in both the RF-Bcl6hi (R2 = 0.32, P < 0.001) and RF-Bcl6int (R2 = 0.14, P < 0.001) groups when individual follicles were analyzed (Figure 3A). Notably, this correlation was stronger and more consistently significant for individual donors in the RF-Bcl6hi group compared with RF-Bcl6int (Supplemental Figure 5, A and B).
 Figure 3
Figure 3Increasing Bcl6 expression is associated with altered spatial distribution of RF immune cell subsets. (A) Correlation between PD1hiCD57hi Tfh cells and CD20hi/dimKi67hiBcl6hi B cells across follicles in LD-LNs. Left panel: RF-Bcl6int donors (pink). Right panel: RF-Bcl6hi donors (green). R² and P values are reported. (B) Ratio of CD20hi/dimKi67hiBcl6hi B cells to PD1hiCD57hi Tfh cells in RF-Bcl6int (pink) versus RF-Bcl6hi (green) donors. (C) Net area under the G-function curve (AUCabove – AUCbelow) by distance (r, μm), comparing spatial organization of GC B cells and Tfh cells subsets across groups (tissues from CHUV were analyzed). (D) Left panel: Connectivity plot from Donor 17-Follicle 6 (RF-Bcl6hi) showing spatial links from the most central GC B cell (yellow) to Tfh cells (green). Right panel: Histogram of minimum Euclidean distances from Donor 17-Follicle 6 (RF-Bcl6hi). (E) Dot plot of mean minimum Euclidean distances between CD20hi/dimKi67hi B cells and PD1hiCD57hi Tfh cells in HLN-Controls (blue), RF-Bcl6int (pink), and RF-Bcl6hi (green). Each point represents one follicle. Mann-Whitney U; *P ≤ 0.05, **P ≤ 0.01, ****P ≤ 0.0001. Raw P-values are shown on the graph, and both raw and FDR-adjusted P-values are reported in Supplemental Table 11.
Furthermore, the ratio of CD20hi/dimKi67hiBcl6hi B cells to PD1hiCD57hi Tfh cells was significantly higher in RF-Bcl6hi compared with RF-Bcl6int (Figure 3B), suggesting an increased likelihood of productive B/T cell interactions in RF-Bcl6hi follicles.
To further investigate the spatial dynamics of GC B cell and Tfh cell interactions, we extracted the spatial coordinates of CD20hi/dimKi67hi, CD20hi/dimKi67hiBcl6hi, PD1hi, and PD1hiCD57hi cells in RFs containing at least 20 cells per phenotype. We then applied the G-function, a spatial statistical method that quantifies cell clustering or dispersion by comparing observed spatial distributions to a random Poisson model using net AUC as a metric (Figure 3C and Supplemental Figure 6A). We found that CD20hi/dimKi67hi and CD20hi/dimKi67hiBcl6hi B cells exhibited a significantly less scattered (more clustered) distribution in RF-Bcl6int tissue compared with RF-Bcl6hi (Figure 3C). In contrast, PD1hi Tfh cells displayed the opposite pattern, with a more dispersed distribution in RF-Bcl6int tissue (Figure 3C). Furthermore, RF-Bcl6hi follicles exhibited a significantly lower clustering profile of both B cells and Tfh cell subsets compared with HLN-Controls of non-COVID-19–infected patients (Figure 3C).
Our distance matrix analysis (an example is shown in Figure 3D) revealed a comparable mean minimum Euclidean distance between proliferating CD20hi/dimKi67hi B cells and PD1hiCD57lo Tfh cells (Supplemental Figure 6B) and between GC B cells CD20hi/dimKi67hiBcl6hi, and PD1hiCD57hi Tfh cells (Supplemental Figure 6B). However, a significantly shorter distance between CD20hi/dimKi67hi B cells and PD1hiCD57hi Tfh cells was observed in RF-Bcl6hi compared with RF-Bcl6int (P = 0.0432) and HLN-Controls (P = 0.0046) (Figure 3E), suggesting a higher possibility for direct B/Tfh interactions in RF-Bcl6hi LD-LNs.
Altogether, our data suggest that increasing Bcl6 expression in RFs is associated with enhanced spatial proximity and interaction between proliferating B cells and highly differentiated Tfh cells.
COVID-19 infection reveals a disconnection in RF immunoreactivity between LD-LNs and matched distal subdiaphragmatic LNs. We analyzed lung-draining LNs and subdiaphragmatic LNs (SD-LNs; serving as “internal controls”) from the same donors (CHUV site) (Figure 4, A–C, Supplemental Table 8, and Supplemental Figure 6C) to investigate whether COVID-19 infection leads to a generalized RF Bcl6 immunoreactivity. The adaptive immunity panel (Supplemental Tables 2 and 3) was applied to SD-LNs (Figure 4A).
 Figure 4
Figure 4COVID-19 infection reveals a disconnection in F/GC immunoreactivity between LD-LNs and matched distal subdiaphragmatic LNs. (A) Representative fluorescence images of follicular areas from subdiaphragmatic and matched LD-LNs in 2 RF-Bcl6hi donors. Upper panel: Donor 21. Lower panel: Donor 17. Staining shows CD20hi (blue), Ki67hi (magenta), Bcl6hi (cyan), along with merged composite images. Scale bars: Original magnification, 100 μm (original); 10 μm (zoomed area). (B) Dot plots showing normalized densities of B cells and CD4 T cells (cells/μm²) in subdiaphragmatic (blue) and matched LD-LNs (red). (C) Dot plots showing frequencies of B cell and CD4 T cell subsets (% of total cells) in subdiaphragmatic (blue) and matched LD-LNs (red). Tissues from CHUV were analyzed. Wilcoxon test; *P ≤ 0.05.
Overall, we observed a tendency toward lower cell densities of B cell and Tfh cell subsets in SD-LNs, with a significant decrease in CD20hi/dimKi67hiBcl6hi cells compared with their LD-LN counterparts (Figure 4B). Consistent with this profile, SD-LNs exhibited significantly lower frequencies of CD20hi/dimKi67hiBcl6hi, PD1hiKi67hi, and PD1hiKi67hiBcl6hi cells compared with LD-LNs (Figure 4C).
Our data suggest that the heightened RF activity seen in LD-LNs possibly represents a localized adaptation in lung-draining lymphoid tissues rather than a generalized systemic response.
The RF-Bcl6hi tissues are characterized by a distinct in situ follicular transcriptomic profile. The GeoMx platform was applied to RF areas covering the 3 groups (CHUV site), using normalized and batch-corrected data for transcriptomic analysis (Supplemental Figure 7, A and B). Principal component analysis (PCA) revealed distinct transcriptional profiles based on Bcl6 expression, suggesting molecular differences in GC reactivity across the groups (Figure 5A).
 Figure 5
Figure 5Distinct in situ transcriptomic profiles are associated with RF-Bcl6hi follicles in LD-LNs. (A) Principal component analysis (PCA) of gene expression following normalization and batch correction. Samples are color-coded by group — RF-Bcl6hi (green), RF-Bcl6int (orange), and RF-Bcl6no/lo (purple). (B) Volcano plot showing differentially expressed genes between RF-Bcl6no/lo and RF-Bcl6hi individuals. Orange dots indicate genes upregulated in RF-Bcl6no/lo, purple dots in RF-Bcl6hi, and gray dots denote nonsignificant genes. (C) Heatmaps of differentially expressed genes (DEGs) between RF-Bcl6no/lo and RF-Bcl6hi individuals. Left panel: DEGs associated with TH1 responses, upregulated in the RF-Bcl6no/lo group. Right panel: DEGs related to germinal center (GC) development, upregulated in the RF-Bcl6hi group. Colors represent Z-scored expression values. (D) Left panel: Volcano plot showing differentially expressed genes (DEGs) between RF-Bcl6int and RF-Bcl6hi individuals. Orange dots indicate genes significantly upregulated in RF-Bcl6int, while purple dots represent those upregulated in RF-Bcl6hi. Right panel: Heatmap displaying DEGs upregulated in RF-Bcl6hi group. Colors indicate standardized Z-scores, reflecting relative gene expression levels across samples. (E) Dot plot of Reactome pathways significantly enriched in RF-Bcl6no/lo versus RF-Bcl6hi individuals. Dot size indicates the number of differentially expressed genes that are upregulated (left column) or downregulated (right column) in RF-Bcl6no/lo. Dot color represents the –log (FDR), corresponding to the negative logarithm of the FDR.
Several differentially expressed genes (DEGs) emerged from the RF-Bcl6no/lo versus RF-Bcl6hi comparison (Figure 5B), highlighting functional divergence in immune pathways. In RF-Bcl6hi donors, genes linked to GC-development and B cell activation (e.g., BCL6, AICDA, IL21R, CXCL13, STAT3, CD79A, MYC, MKI67) were upregulated, reinforcing their role in sustaining GC activity (Figure 5C). Conversely, in RF-Bcl6no/lo donors, genes associated with a TH1-driven immune response and inflammation (e.g., STAT4, TNFRSF10B, TNFRSF1A, CXCR3, TGFB3, IL7, IFNB1, RUNX1) were significantly upregulated, suggesting a shift toward a proinflammatory and interferon-driven environment (Figure 5C).
Analysis of PCA projection of the ROIs based on the selected gene sets revealed a more clustered expression of TH1-related genes in RF-Bcl6no/lo tissues, whereas GC-associated genes exhibited a more scattered profile in RF-Bcl6hi tissues (Supplemental Figure 7C and Supplemental Figure 8A). The data indicate a more homogeneous distribution of TH1-related genes — and presumably of associated signaling pathways — among RFs within a given tissue, in contrast to the more heterogeneous pattern observed for GC-development–related genes.
Comparing RF-Bcl6int versus RF-Bcl6hi tissues (Figure 5D), we found a significant upregulation of genes favoring Tfh/GC formation (e.g., CXCL13, CD22, MIF, STAT3, AICDA) in RF-Bcl6hi tissues (Figure 5D). Examining expression profiles across all groups, GC-promoting genes (e.g., AICDA, BCL6, IL21R, CD19) showed a progressive upregulation from RF-Bcl6no/lo to RF-Bcl6hi (Supplemental Figure 8B). This was further supported by increased expression of CXCL13, S1PR2, and PCNA, key markers of GC organization and proliferation. Meanwhile, TH1-associated genes (e.g., STAT4, TNFRSF10B, TNFRSF1B) were consistently downregulated in RF-Bcl6hi compared with RF-Bcl6no/lo, whereas RF-Bcl6int exhibited an intermediate profile, with some TH1-related genes (e.g., CXCR3, IFNB1) maintaining moderate expression (Supplemental Figure 8B).
Pathway enrichment analysis revealed upregulation of TNF-related (e.g., TNFR1 Signaling, NF-κβ, TRAIL Signaling) and IFN pathways (e.g., α/β Interferon Signaling) in RF-Bcl6no/lo tissues, suggesting a sustained inflammatory response in these follicles (Figure 5E). Conversely, RF-Bcl6hi tissues showed enrichment of pathways related to metabolism, DNA repair, and immune activation (e.g., Respiratory Electron Transport, ATP Synthesis, Mismatch Repair), suggesting enhanced GC maintenance and cellular homeostasis. Furthermore, the RF-Bcl6hi group showed upregulation of RF-development pathways (e.g., IL-4 signaling, IL-6 signaling, detoxification of reactive oxygen species) compared with RF-Bcl6int (Supplemental Figure 8C). Overall, our findings suggest that RF-Bcl6 expression in COVID-19 LD-LNs is linked to distinct in situ molecular profiles.
B cells and Tfh cells differentially influence TNF signaling depending on RF-Bcl6 expression. To further characterize the cellular composition and gene expression profiles in RF-Bcl6hi and RF-Bcl6no/lo tissues, we conducted a cell deconvolution analysis using single-cell RNA-Seq data from HIV LNs as a reference. This approach enabled us to evaluate the relative abundance of immune cell populations across tissue groups. Our analysis examined the relationship between gene expression and the relative proportions of specific cell types (B cells or Tfh cells). The expression of control genes (with known expression profiles) showed a significant positive association between B cells and B cell–related genes (e.g., CD19, CD22, CD74, CD79A) as well as between Tfh cells and (e.g., ICOS), supporting the validity of our pipeline (Figure 6A).
 Figure 6
Figure 6Association between TNF-family gene expression and follicular B cells or Tfh cells. (A) Dot plots illustrating the correlation between the expression levels of selected control genes — chosen for their established cell type specificity — and the inferred percentages of B cells or Tfh cells obtained through transcriptomic cell deconvolution analysis. The y axis displays batch-corrected and normalized gene expression values, allowing for cross-sample comparability. Each data point represents an individual follicle (region of interest, ROI) derived from tissue samples classified as either RF-Bcl6no/lo (green) or RF-Bcl6hi (orange), enabling visualization of expression trends across reactive follicle subtypes. (B) Dot plots showing the correlation between the expression levels of TNF family genes and the inferred proportions of B cells or Tfh cells, as determined by transcriptomic deconvolution. The y axis represents batch-corrected and normalized gene expression values, facilitating robust comparison across samples. Each point corresponds to a distinct follicle (region of interest, ROI) originating from either the RF-Bcl6no/lo group (green) or the RF-Bcl6hi group (orange), allowing assessment of how TNF-related signaling may associate with specific immune cell populations. For each correlation, the coefficient of determination (R²) and corresponding P value are reported, indicating the strength and significance of the observed associations.
We observed heterogeneous associations between B cells, Tfh cells, and TNF-family genes (Figure 6B), which were differentially expressed between RF-Bcl6no/lo and RF-Bcl6hi samples (Figure 6B). In the RF-Bcl6no/lo group, a strong negative association was detected between B cells and several genes (e.g., TNFRSF1A/TNF-R1, TNFRSF1B/TNF-R2, TRAF1, TNFRSF4/CD134, and TNFRSF25/DR3) (Figure 6B). This negative association was less pronounced for Tfh cells (Figure 6B). In contrast, the RF-Bcl6hi group showed a trend toward a positive association between several genes and B cells, reaching significance for TRAF2 (Figure 6B), as well as between Tfh cells and TNFRSF1A/TNF-R1 (Figure 6B).
Together, these findings suggest that differences in TNF-signaling gene expression across RF-Bcl6 groups may reflect distinct immune microenvironments, with altered B cell and Tfh cell dynamics potentially contributing to the divergent follicular responses observed in RF-Bcl6hi versus RF-Bcl6no/lo tissues.
RF-Bcl6no/lo tissues are characterized by a distinct in situ follicular macrophage–associated gene-expression profile. Next, we investigated the prevalence of innate immunity subsets (CD68, CD14 for monocytes/macrophages, and MPO, a marker of granulocytes) for all available tissues (Supplemental Figure 9A). Since the innate antibody panel (Supplemental Table 3) did not include follicular markers, our histocytometry analysis was applied to both F and EF areas (Supplemental Figure 9B). We observed a significant association between CD68 (P = 0.002) (Figure 7A and Supplemental Table 9) or CD14 (P = 0.024) (Figure 7A and Supplemental Table 9). Histocytometry-calculated counts and death postinfection (DPI) in RF-Bcl6no/lo donors (CHUV site), suggesting a link between follicular monocytes/macrophages and disease severity and clinical outcome.
 Figure 7
Figure 7RF-Bcl6no/lo follicles exhibit a distinct in situ macrophage profile in COVID-19 LD-LNs. (A) Left panel: Linear regression analysis of log-transformed CD68 expression versus days post-infection (DPI) in LD-LNs from COVID-19–infected individuals. Right panel: Linear regression analysis of log-transformed CD14 expression versus DPI. Log expression levels of CD68 and CD14 are plotted on the y axis, and DPI on the x axis. Data points represent individual donors: RF-Bcl6no/lo (purple circles), RF-Bcl6int (orange circles), and RF-Bcl6hi (green circles). Regression lines illustrate group-specific trends. Statistical results for CD68 and CD14 are summarized in Supplemental Table 9. (B) Heatmap showing macrophage-specific differentially expressed genes (DEGs) in RF-Bcl6no/lo versus RF-Bcl6hi individuals. The y axis lists macrophage-related genes, and the x axis displays individual donors, grouped by RF-Bcl6hi (left) and RF-Bcl6no/lo (right). Colors represent Z-scored expression values. (C) Multiple linear regression analysis comparing predicted versus observed Bcl6 expression levels. Data points are color-coded by group: RF-Bcl6no/lo (purple), RF-Bcl6int (orange), and RF-Bcl6hi (green). The dashed line represents a perfect correlation between predicted and actual values. Supplemental Table 10 summarizes the regression model, including marker predictors, estimated coefficients, standard errors, t-values, and P values. Significant associations were found for CD20 Ki67 (positive predictor) and CD68 (negative predictor).
To gain further insight into the potential role of monocyte/macrophages, we further investigated macrophage-associated gene expression across tissue groups. Using our GeoMx dataset, we selected genes related to RF macrophage activity (e.g., IL12B, CXCL9, CCL7, CD163L1, and CD68) and compared their expression levels between groups. An overall upregulation of these genes was found in RF-Bcl6no/lo compared with RF-Bcl6hi (Figure 7B), suggesting a more pronounced macrophage-driven inflammatory response in the absence of robust GC formation. To further investigate the link between macrophages and Bcl6 expression, we built a multiple-regression model using total cell counts from our histocytometry data, integrating both adaptive and innate panels. Major immune populations were included (CD20hi/dim, CD20hi/dimKi67hi, PD1hi, PD1hiKi67hi, CD8hi, CD8hiGrzBhi, CD14hi, CD68hi, and MPOhi), excluding those directly involving Bcl6 staining. The model showed high predictive accuracy, with predicted Bcl6 expression closely matching observed levels across RF-Bcl6 groups (Figure 7C). Regression analysis revealed a strong positive association between Bcl6 and CD20hi/dimKi67hi levels (β = 0.124, P = 2.3 × 10–6) and a significant negative association with CD68hi expression (β = –0.002, P = 0.012) (Supplemental Table 10), suggesting that macrophage infiltration may suppress Bcl6 expression. These findings support a model in which elevated macrophage levels in RF-Bcl6no/lo tissues could limit GC responses.











