Research ArticleImmunologyInfectious diseasePublic Health
Open Access | ![]() 10.1172/jci.insight.198045
10.1172/jci.insight.198045
Intranasal booster drives class switching and homing of memory B cells for mucosal IgA response
Si Chen,1,2,3,4 Zhengyuan Zhang,3,5 Zihan Lin,3,5 Li Yin,3,5 Lishan Ning,4 Wenming Liu,3 Qian Wang,1 Chenchen Yang,1,4 Bo Feng,4 Ying Feng,4 Yongping Wang,3,5 Hengchun Li,4 Ping He,4 Huan Liang,1 Yichu Liu,3 Zhixia Li,3 Bo Liu,4 Yang Li,6 Diana Boraschi,6 Linbing Qu,3 Xuefeng Niu,1 Nanshan Zhong,1,4 Pingchao Li,3 and Ling Chen1,2,3,4
1State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, Guangzhou, China.
2Guangzhou Institute of Infectious Disease, Guangzhou Eighth People’s Hospital, Guangzhou Medical University, Guangzhou, China.
3State Key Laboratory of Respiratory Disease, Human Cell Lineage Atlas Facility, Guangdong Provincial Key Laboratory of Biocomputing, Institute of Drug Discovery, Guangzhou Institutes of Biomedicine and Health, Chinese Academy of Sciences, Guangzhou, China.
4Guangzhou National Laboratory, Guangzhou, China.
5University of Chinese Academy of Sciences, Beijing, China.
6Shenzhen Institutes of Advanced Technology, Shenzhen, China.
Address correspondence to: Ling Chen, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.32015289; Email: chen_ling@gibh.ac.cn. Or to: Pingchao Li, State Key Laboratory of Respiratory Disease, Human Cell Lineage Atlas Facility, Guangdong Provincial Key Laboratory of Biocomputing, Institute of Drug Discovery, Guangzhou Institutes of Biomedicine and Health, Chinese Academy of Sciences, 190# Kaiyuan Avenue, huangpu District, Guangzhou 510700, China. Phone: 86.20.32015289; Email: li_pingchao@gibh.ac.cn. Or to: Nanshan Zhong, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.81567941; Email: nanshan@vip.163.com. Or to: Xuefeng Niu, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.32015289; Email: niuxuefeng@gird.cn.
Authorship note: SC, ZZ, Z Lin, and LY contributed equally to this work.
Find articles by Chen, S. in: PubMed | Google Scholar
1State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, Guangzhou, China.
2Guangzhou Institute of Infectious Disease, Guangzhou Eighth People’s Hospital, Guangzhou Medical University, Guangzhou, China.
3State Key Laboratory of Respiratory Disease, Human Cell Lineage Atlas Facility, Guangdong Provincial Key Laboratory of Biocomputing, Institute of Drug Discovery, Guangzhou Institutes of Biomedicine and Health, Chinese Academy of Sciences, Guangzhou, China.
4Guangzhou National Laboratory, Guangzhou, China.
5University of Chinese Academy of Sciences, Beijing, China.
6Shenzhen Institutes of Advanced Technology, Shenzhen, China.
Address correspondence to: Ling Chen, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.32015289; Email: chen_ling@gibh.ac.cn. Or to: Pingchao Li, State Key Laboratory of Respiratory Disease, Human Cell Lineage Atlas Facility, Guangdong Provincial Key Laboratory of Biocomputing, Institute of Drug Discovery, Guangzhou Institutes of Biomedicine and Health, Chinese Academy of Sciences, 190# Kaiyuan Avenue, huangpu District, Guangzhou 510700, China. Phone: 86.20.32015289; Email: li_pingchao@gibh.ac.cn. Or to: Nanshan Zhong, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.81567941; Email: nanshan@vip.163.com. Or to: Xuefeng Niu, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.32015289; Email: niuxuefeng@gird.cn.
Authorship note: SC, ZZ, Z Lin, and LY contributed equally to this work.
Find articles by Zhang, Z. in: PubMed | Google Scholar
1State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, Guangzhou, China.
2Guangzhou Institute of Infectious Disease, Guangzhou Eighth People’s Hospital, Guangzhou Medical University, Guangzhou, China.
3State Key Laboratory of Respiratory Disease, Human Cell Lineage Atlas Facility, Guangdong Provincial Key Laboratory of Biocomputing, Institute of Drug Discovery, Guangzhou Institutes of Biomedicine and Health, Chinese Academy of Sciences, Guangzhou, China.
4Guangzhou National Laboratory, Guangzhou, China.
5University of Chinese Academy of Sciences, Beijing, China.
6Shenzhen Institutes of Advanced Technology, Shenzhen, China.
Address correspondence to: Ling Chen, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.32015289; Email: chen_ling@gibh.ac.cn. Or to: Pingchao Li, State Key Laboratory of Respiratory Disease, Human Cell Lineage Atlas Facility, Guangdong Provincial Key Laboratory of Biocomputing, Institute of Drug Discovery, Guangzhou Institutes of Biomedicine and Health, Chinese Academy of Sciences, 190# Kaiyuan Avenue, huangpu District, Guangzhou 510700, China. Phone: 86.20.32015289; Email: li_pingchao@gibh.ac.cn. Or to: Nanshan Zhong, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.81567941; Email: nanshan@vip.163.com. Or to: Xuefeng Niu, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.32015289; Email: niuxuefeng@gird.cn.
Authorship note: SC, ZZ, Z Lin, and LY contributed equally to this work.
Find articles by Lin, Z. in: PubMed | Google Scholar
1State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, Guangzhou, China.
2Guangzhou Institute of Infectious Disease, Guangzhou Eighth People’s Hospital, Guangzhou Medical University, Guangzhou, China.
3State Key Laboratory of Respiratory Disease, Human Cell Lineage Atlas Facility, Guangdong Provincial Key Laboratory of Biocomputing, Institute of Drug Discovery, Guangzhou Institutes of Biomedicine and Health, Chinese Academy of Sciences, Guangzhou, China.
4Guangzhou National Laboratory, Guangzhou, China.
5University of Chinese Academy of Sciences, Beijing, China.
6Shenzhen Institutes of Advanced Technology, Shenzhen, China.
Address correspondence to: Ling Chen, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.32015289; Email: chen_ling@gibh.ac.cn. Or to: Pingchao Li, State Key Laboratory of Respiratory Disease, Human Cell Lineage Atlas Facility, Guangdong Provincial Key Laboratory of Biocomputing, Institute of Drug Discovery, Guangzhou Institutes of Biomedicine and Health, Chinese Academy of Sciences, 190# Kaiyuan Avenue, huangpu District, Guangzhou 510700, China. Phone: 86.20.32015289; Email: li_pingchao@gibh.ac.cn. Or to: Nanshan Zhong, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.81567941; Email: nanshan@vip.163.com. Or to: Xuefeng Niu, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.32015289; Email: niuxuefeng@gird.cn.
Authorship note: SC, ZZ, Z Lin, and LY contributed equally to this work.
Find articles by Yin, L. in: PubMed | Google Scholar
1State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, Guangzhou, China.
2Guangzhou Institute of Infectious Disease, Guangzhou Eighth People’s Hospital, Guangzhou Medical University, Guangzhou, China.
3State Key Laboratory of Respiratory Disease, Human Cell Lineage Atlas Facility, Guangdong Provincial Key Laboratory of Biocomputing, Institute of Drug Discovery, Guangzhou Institutes of Biomedicine and Health, Chinese Academy of Sciences, Guangzhou, China.
4Guangzhou National Laboratory, Guangzhou, China.
5University of Chinese Academy of Sciences, Beijing, China.
6Shenzhen Institutes of Advanced Technology, Shenzhen, China.
Address correspondence to: Ling Chen, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.32015289; Email: chen_ling@gibh.ac.cn. Or to: Pingchao Li, State Key Laboratory of Respiratory Disease, Human Cell Lineage Atlas Facility, Guangdong Provincial Key Laboratory of Biocomputing, Institute of Drug Discovery, Guangzhou Institutes of Biomedicine and Health, Chinese Academy of Sciences, 190# Kaiyuan Avenue, huangpu District, Guangzhou 510700, China. Phone: 86.20.32015289; Email: li_pingchao@gibh.ac.cn. Or to: Nanshan Zhong, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.81567941; Email: nanshan@vip.163.com. Or to: Xuefeng Niu, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.32015289; Email: niuxuefeng@gird.cn.
Authorship note: SC, ZZ, Z Lin, and LY contributed equally to this work.
Find articles by Ning, L. in: PubMed | Google Scholar
1State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, Guangzhou, China.
2Guangzhou Institute of Infectious Disease, Guangzhou Eighth People’s Hospital, Guangzhou Medical University, Guangzhou, China.
3State Key Laboratory of Respiratory Disease, Human Cell Lineage Atlas Facility, Guangdong Provincial Key Laboratory of Biocomputing, Institute of Drug Discovery, Guangzhou Institutes of Biomedicine and Health, Chinese Academy of Sciences, Guangzhou, China.
4Guangzhou National Laboratory, Guangzhou, China.
5University of Chinese Academy of Sciences, Beijing, China.
6Shenzhen Institutes of Advanced Technology, Shenzhen, China.
Address correspondence to: Ling Chen, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.32015289; Email: chen_ling@gibh.ac.cn. Or to: Pingchao Li, State Key Laboratory of Respiratory Disease, Human Cell Lineage Atlas Facility, Guangdong Provincial Key Laboratory of Biocomputing, Institute of Drug Discovery, Guangzhou Institutes of Biomedicine and Health, Chinese Academy of Sciences, 190# Kaiyuan Avenue, huangpu District, Guangzhou 510700, China. Phone: 86.20.32015289; Email: li_pingchao@gibh.ac.cn. Or to: Nanshan Zhong, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.81567941; Email: nanshan@vip.163.com. Or to: Xuefeng Niu, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.32015289; Email: niuxuefeng@gird.cn.
Authorship note: SC, ZZ, Z Lin, and LY contributed equally to this work.
Find articles by Liu, W. in: PubMed | Google Scholar
1State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, Guangzhou, China.
2Guangzhou Institute of Infectious Disease, Guangzhou Eighth People’s Hospital, Guangzhou Medical University, Guangzhou, China.
3State Key Laboratory of Respiratory Disease, Human Cell Lineage Atlas Facility, Guangdong Provincial Key Laboratory of Biocomputing, Institute of Drug Discovery, Guangzhou Institutes of Biomedicine and Health, Chinese Academy of Sciences, Guangzhou, China.
4Guangzhou National Laboratory, Guangzhou, China.
5University of Chinese Academy of Sciences, Beijing, China.
6Shenzhen Institutes of Advanced Technology, Shenzhen, China.
Address correspondence to: Ling Chen, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.32015289; Email: chen_ling@gibh.ac.cn. Or to: Pingchao Li, State Key Laboratory of Respiratory Disease, Human Cell Lineage Atlas Facility, Guangdong Provincial Key Laboratory of Biocomputing, Institute of Drug Discovery, Guangzhou Institutes of Biomedicine and Health, Chinese Academy of Sciences, 190# Kaiyuan Avenue, huangpu District, Guangzhou 510700, China. Phone: 86.20.32015289; Email: li_pingchao@gibh.ac.cn. Or to: Nanshan Zhong, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.81567941; Email: nanshan@vip.163.com. Or to: Xuefeng Niu, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.32015289; Email: niuxuefeng@gird.cn.
Authorship note: SC, ZZ, Z Lin, and LY contributed equally to this work.
Find articles by Wang, Q. in: PubMed | Google Scholar
1State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, Guangzhou, China.
2Guangzhou Institute of Infectious Disease, Guangzhou Eighth People’s Hospital, Guangzhou Medical University, Guangzhou, China.
3State Key Laboratory of Respiratory Disease, Human Cell Lineage Atlas Facility, Guangdong Provincial Key Laboratory of Biocomputing, Institute of Drug Discovery, Guangzhou Institutes of Biomedicine and Health, Chinese Academy of Sciences, Guangzhou, China.
4Guangzhou National Laboratory, Guangzhou, China.
5University of Chinese Academy of Sciences, Beijing, China.
6Shenzhen Institutes of Advanced Technology, Shenzhen, China.
Address correspondence to: Ling Chen, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.32015289; Email: chen_ling@gibh.ac.cn. Or to: Pingchao Li, State Key Laboratory of Respiratory Disease, Human Cell Lineage Atlas Facility, Guangdong Provincial Key Laboratory of Biocomputing, Institute of Drug Discovery, Guangzhou Institutes of Biomedicine and Health, Chinese Academy of Sciences, 190# Kaiyuan Avenue, huangpu District, Guangzhou 510700, China. Phone: 86.20.32015289; Email: li_pingchao@gibh.ac.cn. Or to: Nanshan Zhong, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.81567941; Email: nanshan@vip.163.com. Or to: Xuefeng Niu, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.32015289; Email: niuxuefeng@gird.cn.
Authorship note: SC, ZZ, Z Lin, and LY contributed equally to this work.
Find articles by Yang, C. in: PubMed | Google Scholar
1State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, Guangzhou, China.
2Guangzhou Institute of Infectious Disease, Guangzhou Eighth People’s Hospital, Guangzhou Medical University, Guangzhou, China.
3State Key Laboratory of Respiratory Disease, Human Cell Lineage Atlas Facility, Guangdong Provincial Key Laboratory of Biocomputing, Institute of Drug Discovery, Guangzhou Institutes of Biomedicine and Health, Chinese Academy of Sciences, Guangzhou, China.
4Guangzhou National Laboratory, Guangzhou, China.
5University of Chinese Academy of Sciences, Beijing, China.
6Shenzhen Institutes of Advanced Technology, Shenzhen, China.
Address correspondence to: Ling Chen, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.32015289; Email: chen_ling@gibh.ac.cn. Or to: Pingchao Li, State Key Laboratory of Respiratory Disease, Human Cell Lineage Atlas Facility, Guangdong Provincial Key Laboratory of Biocomputing, Institute of Drug Discovery, Guangzhou Institutes of Biomedicine and Health, Chinese Academy of Sciences, 190# Kaiyuan Avenue, huangpu District, Guangzhou 510700, China. Phone: 86.20.32015289; Email: li_pingchao@gibh.ac.cn. Or to: Nanshan Zhong, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.81567941; Email: nanshan@vip.163.com. Or to: Xuefeng Niu, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.32015289; Email: niuxuefeng@gird.cn.
Authorship note: SC, ZZ, Z Lin, and LY contributed equally to this work.
Find articles by Feng, B. in: PubMed | Google Scholar
1State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, Guangzhou, China.
2Guangzhou Institute of Infectious Disease, Guangzhou Eighth People’s Hospital, Guangzhou Medical University, Guangzhou, China.
3State Key Laboratory of Respiratory Disease, Human Cell Lineage Atlas Facility, Guangdong Provincial Key Laboratory of Biocomputing, Institute of Drug Discovery, Guangzhou Institutes of Biomedicine and Health, Chinese Academy of Sciences, Guangzhou, China.
4Guangzhou National Laboratory, Guangzhou, China.
5University of Chinese Academy of Sciences, Beijing, China.
6Shenzhen Institutes of Advanced Technology, Shenzhen, China.
Address correspondence to: Ling Chen, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.32015289; Email: chen_ling@gibh.ac.cn. Or to: Pingchao Li, State Key Laboratory of Respiratory Disease, Human Cell Lineage Atlas Facility, Guangdong Provincial Key Laboratory of Biocomputing, Institute of Drug Discovery, Guangzhou Institutes of Biomedicine and Health, Chinese Academy of Sciences, 190# Kaiyuan Avenue, huangpu District, Guangzhou 510700, China. Phone: 86.20.32015289; Email: li_pingchao@gibh.ac.cn. Or to: Nanshan Zhong, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.81567941; Email: nanshan@vip.163.com. Or to: Xuefeng Niu, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.32015289; Email: niuxuefeng@gird.cn.
Authorship note: SC, ZZ, Z Lin, and LY contributed equally to this work.
Find articles by Feng, Y. in: PubMed | Google Scholar
1State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, Guangzhou, China.
2Guangzhou Institute of Infectious Disease, Guangzhou Eighth People’s Hospital, Guangzhou Medical University, Guangzhou, China.
3State Key Laboratory of Respiratory Disease, Human Cell Lineage Atlas Facility, Guangdong Provincial Key Laboratory of Biocomputing, Institute of Drug Discovery, Guangzhou Institutes of Biomedicine and Health, Chinese Academy of Sciences, Guangzhou, China.
4Guangzhou National Laboratory, Guangzhou, China.
5University of Chinese Academy of Sciences, Beijing, China.
6Shenzhen Institutes of Advanced Technology, Shenzhen, China.
Address correspondence to: Ling Chen, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.32015289; Email: chen_ling@gibh.ac.cn. Or to: Pingchao Li, State Key Laboratory of Respiratory Disease, Human Cell Lineage Atlas Facility, Guangdong Provincial Key Laboratory of Biocomputing, Institute of Drug Discovery, Guangzhou Institutes of Biomedicine and Health, Chinese Academy of Sciences, 190# Kaiyuan Avenue, huangpu District, Guangzhou 510700, China. Phone: 86.20.32015289; Email: li_pingchao@gibh.ac.cn. Or to: Nanshan Zhong, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.81567941; Email: nanshan@vip.163.com. Or to: Xuefeng Niu, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.32015289; Email: niuxuefeng@gird.cn.
Authorship note: SC, ZZ, Z Lin, and LY contributed equally to this work.
Find articles by Wang, Y. in: PubMed | Google Scholar
1State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, Guangzhou, China.
2Guangzhou Institute of Infectious Disease, Guangzhou Eighth People’s Hospital, Guangzhou Medical University, Guangzhou, China.
3State Key Laboratory of Respiratory Disease, Human Cell Lineage Atlas Facility, Guangdong Provincial Key Laboratory of Biocomputing, Institute of Drug Discovery, Guangzhou Institutes of Biomedicine and Health, Chinese Academy of Sciences, Guangzhou, China.
4Guangzhou National Laboratory, Guangzhou, China.
5University of Chinese Academy of Sciences, Beijing, China.
6Shenzhen Institutes of Advanced Technology, Shenzhen, China.
Address correspondence to: Ling Chen, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.32015289; Email: chen_ling@gibh.ac.cn. Or to: Pingchao Li, State Key Laboratory of Respiratory Disease, Human Cell Lineage Atlas Facility, Guangdong Provincial Key Laboratory of Biocomputing, Institute of Drug Discovery, Guangzhou Institutes of Biomedicine and Health, Chinese Academy of Sciences, 190# Kaiyuan Avenue, huangpu District, Guangzhou 510700, China. Phone: 86.20.32015289; Email: li_pingchao@gibh.ac.cn. Or to: Nanshan Zhong, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.81567941; Email: nanshan@vip.163.com. Or to: Xuefeng Niu, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.32015289; Email: niuxuefeng@gird.cn.
Authorship note: SC, ZZ, Z Lin, and LY contributed equally to this work.
Find articles by Li, H. in: PubMed | Google Scholar
1State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, Guangzhou, China.
2Guangzhou Institute of Infectious Disease, Guangzhou Eighth People’s Hospital, Guangzhou Medical University, Guangzhou, China.
3State Key Laboratory of Respiratory Disease, Human Cell Lineage Atlas Facility, Guangdong Provincial Key Laboratory of Biocomputing, Institute of Drug Discovery, Guangzhou Institutes of Biomedicine and Health, Chinese Academy of Sciences, Guangzhou, China.
4Guangzhou National Laboratory, Guangzhou, China.
5University of Chinese Academy of Sciences, Beijing, China.
6Shenzhen Institutes of Advanced Technology, Shenzhen, China.
Address correspondence to: Ling Chen, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.32015289; Email: chen_ling@gibh.ac.cn. Or to: Pingchao Li, State Key Laboratory of Respiratory Disease, Human Cell Lineage Atlas Facility, Guangdong Provincial Key Laboratory of Biocomputing, Institute of Drug Discovery, Guangzhou Institutes of Biomedicine and Health, Chinese Academy of Sciences, 190# Kaiyuan Avenue, huangpu District, Guangzhou 510700, China. Phone: 86.20.32015289; Email: li_pingchao@gibh.ac.cn. Or to: Nanshan Zhong, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.81567941; Email: nanshan@vip.163.com. Or to: Xuefeng Niu, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.32015289; Email: niuxuefeng@gird.cn.
Authorship note: SC, ZZ, Z Lin, and LY contributed equally to this work.
Find articles by He, P. in: PubMed | Google Scholar
1State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, Guangzhou, China.
2Guangzhou Institute of Infectious Disease, Guangzhou Eighth People’s Hospital, Guangzhou Medical University, Guangzhou, China.
3State Key Laboratory of Respiratory Disease, Human Cell Lineage Atlas Facility, Guangdong Provincial Key Laboratory of Biocomputing, Institute of Drug Discovery, Guangzhou Institutes of Biomedicine and Health, Chinese Academy of Sciences, Guangzhou, China.
4Guangzhou National Laboratory, Guangzhou, China.
5University of Chinese Academy of Sciences, Beijing, China.
6Shenzhen Institutes of Advanced Technology, Shenzhen, China.
Address correspondence to: Ling Chen, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.32015289; Email: chen_ling@gibh.ac.cn. Or to: Pingchao Li, State Key Laboratory of Respiratory Disease, Human Cell Lineage Atlas Facility, Guangdong Provincial Key Laboratory of Biocomputing, Institute of Drug Discovery, Guangzhou Institutes of Biomedicine and Health, Chinese Academy of Sciences, 190# Kaiyuan Avenue, huangpu District, Guangzhou 510700, China. Phone: 86.20.32015289; Email: li_pingchao@gibh.ac.cn. Or to: Nanshan Zhong, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.81567941; Email: nanshan@vip.163.com. Or to: Xuefeng Niu, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.32015289; Email: niuxuefeng@gird.cn.
Authorship note: SC, ZZ, Z Lin, and LY contributed equally to this work.
Find articles by Liang, H. in: PubMed | Google Scholar
1State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, Guangzhou, China.
2Guangzhou Institute of Infectious Disease, Guangzhou Eighth People’s Hospital, Guangzhou Medical University, Guangzhou, China.
3State Key Laboratory of Respiratory Disease, Human Cell Lineage Atlas Facility, Guangdong Provincial Key Laboratory of Biocomputing, Institute of Drug Discovery, Guangzhou Institutes of Biomedicine and Health, Chinese Academy of Sciences, Guangzhou, China.
4Guangzhou National Laboratory, Guangzhou, China.
5University of Chinese Academy of Sciences, Beijing, China.
6Shenzhen Institutes of Advanced Technology, Shenzhen, China.
Address correspondence to: Ling Chen, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.32015289; Email: chen_ling@gibh.ac.cn. Or to: Pingchao Li, State Key Laboratory of Respiratory Disease, Human Cell Lineage Atlas Facility, Guangdong Provincial Key Laboratory of Biocomputing, Institute of Drug Discovery, Guangzhou Institutes of Biomedicine and Health, Chinese Academy of Sciences, 190# Kaiyuan Avenue, huangpu District, Guangzhou 510700, China. Phone: 86.20.32015289; Email: li_pingchao@gibh.ac.cn. Or to: Nanshan Zhong, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.81567941; Email: nanshan@vip.163.com. Or to: Xuefeng Niu, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.32015289; Email: niuxuefeng@gird.cn.
Authorship note: SC, ZZ, Z Lin, and LY contributed equally to this work.
Find articles by Liu, Y. in: PubMed | Google Scholar
1State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, Guangzhou, China.
2Guangzhou Institute of Infectious Disease, Guangzhou Eighth People’s Hospital, Guangzhou Medical University, Guangzhou, China.
3State Key Laboratory of Respiratory Disease, Human Cell Lineage Atlas Facility, Guangdong Provincial Key Laboratory of Biocomputing, Institute of Drug Discovery, Guangzhou Institutes of Biomedicine and Health, Chinese Academy of Sciences, Guangzhou, China.
4Guangzhou National Laboratory, Guangzhou, China.
5University of Chinese Academy of Sciences, Beijing, China.
6Shenzhen Institutes of Advanced Technology, Shenzhen, China.
Address correspondence to: Ling Chen, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.32015289; Email: chen_ling@gibh.ac.cn. Or to: Pingchao Li, State Key Laboratory of Respiratory Disease, Human Cell Lineage Atlas Facility, Guangdong Provincial Key Laboratory of Biocomputing, Institute of Drug Discovery, Guangzhou Institutes of Biomedicine and Health, Chinese Academy of Sciences, 190# Kaiyuan Avenue, huangpu District, Guangzhou 510700, China. Phone: 86.20.32015289; Email: li_pingchao@gibh.ac.cn. Or to: Nanshan Zhong, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.81567941; Email: nanshan@vip.163.com. Or to: Xuefeng Niu, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.32015289; Email: niuxuefeng@gird.cn.
Authorship note: SC, ZZ, Z Lin, and LY contributed equally to this work.
Find articles by Li, Z. in: PubMed | Google Scholar
1State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, Guangzhou, China.
2Guangzhou Institute of Infectious Disease, Guangzhou Eighth People’s Hospital, Guangzhou Medical University, Guangzhou, China.
3State Key Laboratory of Respiratory Disease, Human Cell Lineage Atlas Facility, Guangdong Provincial Key Laboratory of Biocomputing, Institute of Drug Discovery, Guangzhou Institutes of Biomedicine and Health, Chinese Academy of Sciences, Guangzhou, China.
4Guangzhou National Laboratory, Guangzhou, China.
5University of Chinese Academy of Sciences, Beijing, China.
6Shenzhen Institutes of Advanced Technology, Shenzhen, China.
Address correspondence to: Ling Chen, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.32015289; Email: chen_ling@gibh.ac.cn. Or to: Pingchao Li, State Key Laboratory of Respiratory Disease, Human Cell Lineage Atlas Facility, Guangdong Provincial Key Laboratory of Biocomputing, Institute of Drug Discovery, Guangzhou Institutes of Biomedicine and Health, Chinese Academy of Sciences, 190# Kaiyuan Avenue, huangpu District, Guangzhou 510700, China. Phone: 86.20.32015289; Email: li_pingchao@gibh.ac.cn. Or to: Nanshan Zhong, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.81567941; Email: nanshan@vip.163.com. Or to: Xuefeng Niu, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.32015289; Email: niuxuefeng@gird.cn.
Authorship note: SC, ZZ, Z Lin, and LY contributed equally to this work.
Find articles by Liu, B. in: PubMed | Google Scholar
1State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, Guangzhou, China.
2Guangzhou Institute of Infectious Disease, Guangzhou Eighth People’s Hospital, Guangzhou Medical University, Guangzhou, China.
3State Key Laboratory of Respiratory Disease, Human Cell Lineage Atlas Facility, Guangdong Provincial Key Laboratory of Biocomputing, Institute of Drug Discovery, Guangzhou Institutes of Biomedicine and Health, Chinese Academy of Sciences, Guangzhou, China.
4Guangzhou National Laboratory, Guangzhou, China.
5University of Chinese Academy of Sciences, Beijing, China.
6Shenzhen Institutes of Advanced Technology, Shenzhen, China.
Address correspondence to: Ling Chen, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.32015289; Email: chen_ling@gibh.ac.cn. Or to: Pingchao Li, State Key Laboratory of Respiratory Disease, Human Cell Lineage Atlas Facility, Guangdong Provincial Key Laboratory of Biocomputing, Institute of Drug Discovery, Guangzhou Institutes of Biomedicine and Health, Chinese Academy of Sciences, 190# Kaiyuan Avenue, huangpu District, Guangzhou 510700, China. Phone: 86.20.32015289; Email: li_pingchao@gibh.ac.cn. Or to: Nanshan Zhong, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.81567941; Email: nanshan@vip.163.com. Or to: Xuefeng Niu, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.32015289; Email: niuxuefeng@gird.cn.
Authorship note: SC, ZZ, Z Lin, and LY contributed equally to this work.
Find articles by
Li, Y.
in:
PubMed
|
Google Scholar
|

1State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, Guangzhou, China.
2Guangzhou Institute of Infectious Disease, Guangzhou Eighth People’s Hospital, Guangzhou Medical University, Guangzhou, China.
3State Key Laboratory of Respiratory Disease, Human Cell Lineage Atlas Facility, Guangdong Provincial Key Laboratory of Biocomputing, Institute of Drug Discovery, Guangzhou Institutes of Biomedicine and Health, Chinese Academy of Sciences, Guangzhou, China.
4Guangzhou National Laboratory, Guangzhou, China.
5University of Chinese Academy of Sciences, Beijing, China.
6Shenzhen Institutes of Advanced Technology, Shenzhen, China.
Address correspondence to: Ling Chen, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.32015289; Email: chen_ling@gibh.ac.cn. Or to: Pingchao Li, State Key Laboratory of Respiratory Disease, Human Cell Lineage Atlas Facility, Guangdong Provincial Key Laboratory of Biocomputing, Institute of Drug Discovery, Guangzhou Institutes of Biomedicine and Health, Chinese Academy of Sciences, 190# Kaiyuan Avenue, huangpu District, Guangzhou 510700, China. Phone: 86.20.32015289; Email: li_pingchao@gibh.ac.cn. Or to: Nanshan Zhong, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.81567941; Email: nanshan@vip.163.com. Or to: Xuefeng Niu, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.32015289; Email: niuxuefeng@gird.cn.
Authorship note: SC, ZZ, Z Lin, and LY contributed equally to this work.
Find articles by Boraschi, D. in: PubMed | Google Scholar
1State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, Guangzhou, China.
2Guangzhou Institute of Infectious Disease, Guangzhou Eighth People’s Hospital, Guangzhou Medical University, Guangzhou, China.
3State Key Laboratory of Respiratory Disease, Human Cell Lineage Atlas Facility, Guangdong Provincial Key Laboratory of Biocomputing, Institute of Drug Discovery, Guangzhou Institutes of Biomedicine and Health, Chinese Academy of Sciences, Guangzhou, China.
4Guangzhou National Laboratory, Guangzhou, China.
5University of Chinese Academy of Sciences, Beijing, China.
6Shenzhen Institutes of Advanced Technology, Shenzhen, China.
Address correspondence to: Ling Chen, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.32015289; Email: chen_ling@gibh.ac.cn. Or to: Pingchao Li, State Key Laboratory of Respiratory Disease, Human Cell Lineage Atlas Facility, Guangdong Provincial Key Laboratory of Biocomputing, Institute of Drug Discovery, Guangzhou Institutes of Biomedicine and Health, Chinese Academy of Sciences, 190# Kaiyuan Avenue, huangpu District, Guangzhou 510700, China. Phone: 86.20.32015289; Email: li_pingchao@gibh.ac.cn. Or to: Nanshan Zhong, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.81567941; Email: nanshan@vip.163.com. Or to: Xuefeng Niu, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.32015289; Email: niuxuefeng@gird.cn.
Authorship note: SC, ZZ, Z Lin, and LY contributed equally to this work.
Find articles by
Qu, L.
in:
PubMed
|
Google Scholar
|
1State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, Guangzhou, China.
2Guangzhou Institute of Infectious Disease, Guangzhou Eighth People’s Hospital, Guangzhou Medical University, Guangzhou, China.
3State Key Laboratory of Respiratory Disease, Human Cell Lineage Atlas Facility, Guangdong Provincial Key Laboratory of Biocomputing, Institute of Drug Discovery, Guangzhou Institutes of Biomedicine and Health, Chinese Academy of Sciences, Guangzhou, China.
4Guangzhou National Laboratory, Guangzhou, China.
5University of Chinese Academy of Sciences, Beijing, China.
6Shenzhen Institutes of Advanced Technology, Shenzhen, China.
Address correspondence to: Ling Chen, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.32015289; Email: chen_ling@gibh.ac.cn. Or to: Pingchao Li, State Key Laboratory of Respiratory Disease, Human Cell Lineage Atlas Facility, Guangdong Provincial Key Laboratory of Biocomputing, Institute of Drug Discovery, Guangzhou Institutes of Biomedicine and Health, Chinese Academy of Sciences, 190# Kaiyuan Avenue, huangpu District, Guangzhou 510700, China. Phone: 86.20.32015289; Email: li_pingchao@gibh.ac.cn. Or to: Nanshan Zhong, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.81567941; Email: nanshan@vip.163.com. Or to: Xuefeng Niu, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.32015289; Email: niuxuefeng@gird.cn.
Authorship note: SC, ZZ, Z Lin, and LY contributed equally to this work.
Find articles by Niu, X. in: PubMed | Google Scholar
1State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, Guangzhou, China.
2Guangzhou Institute of Infectious Disease, Guangzhou Eighth People’s Hospital, Guangzhou Medical University, Guangzhou, China.
3State Key Laboratory of Respiratory Disease, Human Cell Lineage Atlas Facility, Guangdong Provincial Key Laboratory of Biocomputing, Institute of Drug Discovery, Guangzhou Institutes of Biomedicine and Health, Chinese Academy of Sciences, Guangzhou, China.
4Guangzhou National Laboratory, Guangzhou, China.
5University of Chinese Academy of Sciences, Beijing, China.
6Shenzhen Institutes of Advanced Technology, Shenzhen, China.
Address correspondence to: Ling Chen, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.32015289; Email: chen_ling@gibh.ac.cn. Or to: Pingchao Li, State Key Laboratory of Respiratory Disease, Human Cell Lineage Atlas Facility, Guangdong Provincial Key Laboratory of Biocomputing, Institute of Drug Discovery, Guangzhou Institutes of Biomedicine and Health, Chinese Academy of Sciences, 190# Kaiyuan Avenue, huangpu District, Guangzhou 510700, China. Phone: 86.20.32015289; Email: li_pingchao@gibh.ac.cn. Or to: Nanshan Zhong, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.81567941; Email: nanshan@vip.163.com. Or to: Xuefeng Niu, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.32015289; Email: niuxuefeng@gird.cn.
Authorship note: SC, ZZ, Z Lin, and LY contributed equally to this work.
Find articles by Zhong, N. in: PubMed | Google Scholar
1State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, Guangzhou, China.
2Guangzhou Institute of Infectious Disease, Guangzhou Eighth People’s Hospital, Guangzhou Medical University, Guangzhou, China.
3State Key Laboratory of Respiratory Disease, Human Cell Lineage Atlas Facility, Guangdong Provincial Key Laboratory of Biocomputing, Institute of Drug Discovery, Guangzhou Institutes of Biomedicine and Health, Chinese Academy of Sciences, Guangzhou, China.
4Guangzhou National Laboratory, Guangzhou, China.
5University of Chinese Academy of Sciences, Beijing, China.
6Shenzhen Institutes of Advanced Technology, Shenzhen, China.
Address correspondence to: Ling Chen, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.32015289; Email: chen_ling@gibh.ac.cn. Or to: Pingchao Li, State Key Laboratory of Respiratory Disease, Human Cell Lineage Atlas Facility, Guangdong Provincial Key Laboratory of Biocomputing, Institute of Drug Discovery, Guangzhou Institutes of Biomedicine and Health, Chinese Academy of Sciences, 190# Kaiyuan Avenue, huangpu District, Guangzhou 510700, China. Phone: 86.20.32015289; Email: li_pingchao@gibh.ac.cn. Or to: Nanshan Zhong, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.81567941; Email: nanshan@vip.163.com. Or to: Xuefeng Niu, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.32015289; Email: niuxuefeng@gird.cn.
Authorship note: SC, ZZ, Z Lin, and LY contributed equally to this work.
Find articles by Li, P. in: PubMed | Google Scholar
1State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, Guangzhou, China.
2Guangzhou Institute of Infectious Disease, Guangzhou Eighth People’s Hospital, Guangzhou Medical University, Guangzhou, China.
3State Key Laboratory of Respiratory Disease, Human Cell Lineage Atlas Facility, Guangdong Provincial Key Laboratory of Biocomputing, Institute of Drug Discovery, Guangzhou Institutes of Biomedicine and Health, Chinese Academy of Sciences, Guangzhou, China.
4Guangzhou National Laboratory, Guangzhou, China.
5University of Chinese Academy of Sciences, Beijing, China.
6Shenzhen Institutes of Advanced Technology, Shenzhen, China.
Address correspondence to: Ling Chen, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.32015289; Email: chen_ling@gibh.ac.cn. Or to: Pingchao Li, State Key Laboratory of Respiratory Disease, Human Cell Lineage Atlas Facility, Guangdong Provincial Key Laboratory of Biocomputing, Institute of Drug Discovery, Guangzhou Institutes of Biomedicine and Health, Chinese Academy of Sciences, 190# Kaiyuan Avenue, huangpu District, Guangzhou 510700, China. Phone: 86.20.32015289; Email: li_pingchao@gibh.ac.cn. Or to: Nanshan Zhong, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.81567941; Email: nanshan@vip.163.com. Or to: Xuefeng Niu, State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, the First Affiliated Hospital of Guangzhou Medical University, 28# Qiaozhong Middle Road, Liwan District, Guangzhou 510120, China. Phone: 86.20.32015289; Email: niuxuefeng@gird.cn.
Authorship note: SC, ZZ, Z Lin, and LY contributed equally to this work.
Find articles by
Chen, L.
in:
PubMed
|
Google Scholar
|
Published December 23, 2025 - More info
JCI Insight. 2026;11(3):e198045. https://doi.org/10.1172/jci.insight.198045.
© 2025 Chen et al. This work is licensed under the Creative Commons Attribution 4.0 International License. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
Received: July 16, 2025; Accepted: December 12, 2025

-
Results
Intranasal booster-induced mucosal sIgA could be up to 100-fold more potent than serum IgG and IgA in neutralizing SARS-CoV-2 variants. All donors previously received 2 injections of inactivated whole-virus vaccine more than 6 months ago and have never been infected with SARS-CoV-2 (Supplemental Table 1; supplemental material available online with this article; https://doi.org/10.1172/jci.insight.198045DS1). There were no substantial nasal spike-specific IgA before intranasal vaccination. Nasal lavages and serum samples from each donor were collected within 3–6 weeks after 2 doses of intranasal spray with Ad5-Omicron spike vaccine, an adenovirus type 5 carrying Omicron BA.1 spike (Supplemental Table 1). Paired nasal sIgA, serum IgG, and serum IgA from the same individuals were purified using affinity chromatography. The average yield of purified nasal sIgA from the nasal lavages of the 6 donors was 1.36 ± 0.31 μg/mL, while the average yields from serum were 9.13 ± 2.38 mg/mL for IgG and 0.75 ± 0.15 mg/mL for IgA (Supplemental Table 7). SDS-PAGE and Western blot analysis confirmed that nasal sIgA from all donors are dimeric and are polymeric forms with molecular weight 400 kD or higher (Supplemental Figure 1). In contrast, serum IgG and IgA are monomeric with molecular weight 150 kD and 160 kD, respectively (Supplemental Figure 1). Nasal sIgA was compared with the same person’s serum IgG and IgA for their neutralizing and spike binding activities. On average, nasal sIgA was 17-, 30-, 125-, and 813-fold more potent than serum IgG in neutralizing SARS-CoV-2 WT, BA.1, BA.5, and XBB.1.5 pseudoviruses. Similarly, nasal sIgA was 61, 49, 95, and 167-fold more potent than serum IgA in neutralizing WT, BA.1, BA.5, and XBB.1.5 (Figure 1, A and B, and Supplemental Tables 2 and 3). For each donor, nasal sIgA was 12- to 37-fold stronger than serum IgG and was 34- to 97-fold stronger than serum IgA in binding to spike proteins of WT, BA.1, BA.5, and XBB.1.5, respectively (Figure 1, C and D, Supplemental Tables 2 and 3). We also accessed the percentage of spike-specific antibodies in the purified sIgA, serum IgA, and serum IgG using mAb 719-1, a broadly neutralizing mAbs (this study) in its respective isotypes as a reference standard. Among these paired nasal lavage and serum samples, there was an average of 0.83% (0.11%–2.38%), 0.59% (0.08%–1.34%), 0.25% (0.03%–0.53%), and 0.20% (0.02%–0.52%) of WT, BA.1, BA.5, and XBB.1.5 spike-specific sIgA in total nasal sIgA. There was an average of 0.33% (0.13%–0.68%), 0.37% (0.15%–0.74%), 0.17% (0.11%–0.34%), and 0.13% (0.07%–0.19%) of WT, BA.1, BA.5, and XBB.1.5 spike-specific IgG in total serum IgG. There was an average of 0.13% (0.02%–0.28%), 0.16% (0.02%–0.40%), 0.05% (0.01%–0.14%), and 0.04% (0.01%–0.07%) of WT, BA.1, BA.5, and XBB.1.5 spike-specific IgA in total serum IgA (Supplemental Figure 5A). We then compared the neutralizing activity of nasal sIgA, serum IgA, and serum IgG based on equal amounts of spike-specific antibodies for each paired antibody isotypes. On average, nasal sIgA exhibited 10-, 23-, 85-, and 2,162-fold greater potency than serum IgG in neutralizing SARS-CoV-2 WT, BA.1, BA.5, and XBB.1.5 pseudoviruses, respectively. Nasal sIgA exhibited 10-, 10-, 17-, and 720-fold more potent than serum IgA in neutralizing WT, BA.1, BA.5, and XBB.1.5 pseudoviruses, respectively (Supplemental Figure 5, B and C). Spearman correlation analysis showed that nasal sIgA neutralizing activity was highly correlated with sIgA spike binding activity (r = 0.8200, P < 0.001), but not with serum IgG neutralizing activity (r = 0.4262, P = 0.078) (Figure 1, E and F). Therefore, nasal sIgA is much more potent than serum IgG and IgA in neutralizing and binding activities. The disparity between nasal sIgA and serum IgG or serum IgA was even more dramatic for lately emerged Omicron XBB.1.5, demonstrating nasal sIgA has enhanced broadly neutralizing breadth than serum antibodies in neutralizing all early and late SARS-CoV-2 variants that we tested.
 Figure 1
Figure 1Neutralizing and spike-binding activities of purified nasal sIgA, serum IgG, and IgA. (A) Neutralizing activities of 6 paired purified nasal sIgA, serum IgG, and serum IgA samples against WT, BA.1, BA.5, and XBB.1.5 were assessed using a lentivirus-based pseudovirus system. The results are presented as 50% inhibitory concentration (IC50) in nM. Samples showing no detectable neutralization activity at the highest concentration (1,000 μg/mL) were assigned an IC50 value of 6,666.7 nM (dashed line), representing no neutralization. Geomean values (n = 6) are reported above the graph. (B) The ratios of IC50 between paired IgG/sIgA, IgA/sIgA, and IgG/IgA for each donor are presented. Mean values (n = 6) are reported above the graph. (C) Binding activities of 6 paired purified nasal sIgA, serum IgG and serum IgA samples against WT, BA.1, BA.5, and XBB.1.5 spike proteins were measured by ELISA. Results are presented as 50% effective concentration (EC50) in nM. Data are calculated as geomean (n = 6) and reported above the graph. (D) The ratios of EC50 between paired IgG/sIgA, IgA/sIgA, and IgG/IgA for each donor are presented. Mean values (n = 6) are reported above the graph. (E and F) Correlation analysis was performed between: nasal sIgA neutralization activity (IC50) and binding activity (EC50) (E), nasal sIgA and serum IgG neutralization activity (IC50) (F). Statistical significance was determined using Spearman’s rank correlation (coefficients and P values are shown).
Obtaining mucosa–associated sIgA mAbs using a combination of mass spectrometry and B cell sequencing. Since few studies have focused on the isolation and characterization of mAbs from nasal mucosa in humans, we combined liquid chromatography–tandem mass spectrometry (LC-MS/MS) for peptide identification of spike-binding sIgA (MS Ig-seq) with single-cell BCR sequencing (scBCR-seq) to obtain paired heavy- and light-chain sequences, which were then cloned into expression plasmids for recombinant production of mucosa-associated sIgA mAbs (Figure 2A). Spike-specific sIgA was purified from the nasal lavage of a donor (Donor 1) at 4 weeks after the second intranasal booster and subsequently analyzed by LC-MS/MS. Peptide identification from the raw LC-MS/MS data was performed using Proteome Discoverer and PEAKS Studio software, which requires a comprehensive antibody database for the search. Accordingly, we constructed a personalized antibody repertoire by performing scBCR-seq on peripheral blood mononuclear cell–derived (PBMC-derived) B cells from the same donor after each vaccination (Supplemental Table 4). A total of 33,675 BCR sequences and 13,433 peptide sequences were obtained (Supplemental Table 4). Alignment of sIgA peptide sequences with BCR immunoglobulin heavy chain (IgH) sequences led to the identification of 45 heavy-chain IgH sequences and their corresponding immunoglobulin light chain (IgL) sequences (Figure 2B and Supplemental Figure 2B). These 45 mucosal mAbs are contributed by 24 IgH germline genes, including IGHV3-23 (7 of 45, 15.6%), IGHV4-39 (4 of 45, 8.9%), IGHV1-18, IGHV3-53, and IGHV4-59 (3 of 45, 6.7%), and by 23 IgL germline genes, including IGKV3-15 (6 of 45, 13.3%), IGKV3-20 (5 of 45, 11.1%), IGKV1-39, IGKV3-11, IGKV4-1, IGLV2-23, and IGLV3-21 (3 of 45, 6.7%) (Figure 2B). The cDNAs were then synthesized and cloned into expression plasmids for production of mAbs in different isotypes. Of the 42 mAbs that were successfully expressed and purified in IgG1 form, 30 mAbs showed strong binding activity to the BA.1 and BA.5 spike proteins, while 12 mAbs showed weak binding activity. Through ELISA, we found that the successfully expressed mAbs bind to either the receptor-binding domain (RBD) or the N-terminal domain (NTD) (Figure 2C and Supplemental Table 5).
 Figure 2
Figure 2Identification of mucosal sIgA mAbs using MS Ig-seq and scBCR-seq. (A) Scheme of the study. The schematic was drawn using BioRender. For MS Ig-seq, BA.1 spike-specific sIgA from a donor was isolated using magnetic beads coupled with BA.1 spike, digested with trypsin, and analyzed using LC-MS/MS. A personalized antibody repertoire was constructed by scBCR-seq from the same donor. cDNAs encoding the heavy chain and light chain of respective antibodies were synthesized and cloned into expression plasmids. (B) The germline heavy- and light-chain gene usage (IGHV, IGKV, and IGLV) and their pairing combinations among the 45 isolated antibodies were visualized using a chord diagram. Each arc represents a heavy-chain and light-chain germline gene pair, the width of which is proportional to the corresponding antibody count. (C) Binding of mucosal mAbs (719-1 to 45, IgG) to variant spike proteins (BA.1, BA.5, XBB.1.5, and JN.1) was measured by ELISA. The chart shows mAbs with EC50 below 5 nM against at least 2 variants. (D) BLI-based competition assays were performed between RBD-binding mAbs 719-1, 719-14, 719-37, 719-40, 719-42, and known antibodies targeting distinct RBD epitopes (LY-CoV-555, P2B-2F6, FC08, S309 and CR3022) or ACE2. The competition percent between each isolated mAb and the known antibodies or ACE2 is presented in the table. (E) Assessment of the neutralizing potency of mAbs 719-1, 719-3, 719-4, 719-14, 719-19, 719-37, 719-39, 719-40, and 719-42 expressed in IgG, mIgA, dIgA, and sIgA forms against pseudoviruses expressing spike from WT and Omicron subvariants BA.1, BA.5, XBB.1.5, JN.1, and KP.3. Data are shown as IC50.
Nasal dIgA and sIgA mAbs have more potent neutralizing and spike-binding activities than their monomeric IgG and IgA isoforms. Nine mAbs with strong neutralizing and spike-binding activities against BA.5 were selected for further study (Figure 2C and Table 1). mAbs 719-3, 719-4, 719-19, and 719-39 bind to the NTD, whereas 719-1, 719-14, 719-37, 719-40, and 719-42 bind to RBD. mAbs 719-14, 719-19, 719-37, 719-39, 719-40, and 719-42 bind to spike proteins of BA.1, BA.5, XBB.1.5, and JN.1. 719-1 and 719-4 bind to spike proteins of BA.1, BA.5, and XBB.1.5. mAb 719-3 binds to spike proteins of BA.1 and BA.5 (Figure 2C). Using biolayer interferometry (BLI) analysis, we found that 719-1, 719-14, and 719-40 competed for RBD binding with well-known RBD III type mAbs FC08 or S309 (20, 21). mAb 719-14 also competes with LY-CoV-555 and P2B-2F6, 2 well-known RBD II type mAbs (22, 23). mAbs 719-37 and 719-42 compete with CR3022, a well-known RBD IV type mAb (24). mAb 719-42 also showed competition with angiotensin-converting enzyme 2 (ACE2) for binding to spike, suggesting its epitope overlaps with ACE2 binding site (Figure 2D). Therefore, nasal mucosal neutralizing mAbs can target a diverse range of epitopes on spike protein.
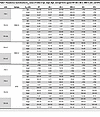 Table 1
Table 1Pseudovirus neutralization IC50 values of mAbs in IgG, mIgA, dIgA, and sIgA forms against WT, BA.1, BA.5, XBB.1.5, JN.1, and KP.3
We next expressed and purified each of these neutralizing mAbs in 4 isotypes — IgG1, mIgA1, dIgA1, and sIgA1 — and confirmed their molecular weights and assessed purities using SDS-PAGE and Western blot analysis (Supplemental Figure 2A). All sIgA isotypes exhibited greater neutralizing potency than the corresponding IgG or mIgA isotypes against at least 1 SARS-CoV-2 variant, with an average enhancement of 13-fold (Figure 2E and Table 1 and Table 2). Some mAbs in sIgA isotype, such as 719-14 and 719-40, exhibited IC50 in subpicomolar range against lately emerged Omicron JN.1 and KP.3. 719-14 sIgA was 58-fold more potent, and 719-40 sIgA was 15-fold more potent than their IgG isotypes in neutralizing KP.3. Interestingly, 719-19 and 719-39 IgG isotypes showed no neutralizing activities against BA.1 and KP.3 (IC50 > 750 nM), but their sIgA isotypes showed an IC50 of 3.42 nM and 23.63 nM against BA.1, and 41.57 nM and 149.45 nM against KP.3, respectively (Figure 2E and Tables 1 and 2). Some sIgA, such as 719-19 sIgA, exhibited 23-fold enhanced neutralizing potency than dIgA against BA.1, demonstrating secretory component (SC) may have marked enhancement on some sIgA mAbs. Further investigation is required to understand how the addition of SC enhances the neutralizing potency of dIgA. Most mAbs in IgG isotype showed similar neutralizing potency as their mIgA isotypes (Figure 2E and Table 1). We also assessed these mAbs in sIgA, dIgA, mIgA, and IgG isotypes for binding activities with spike proteins of WT, BA.1, BA.5, XBB.1.5, JN.1, and KP.3. Most mAbs in sIgA isotype were 2- to 11-fold more potent than their mIgA and IgG isotypes in binding to at least 1 spike protein (Supplemental Figure 3 and Table 3).
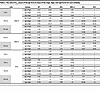 Table 2
Table 2The ratio of IC50 values of the IgG form to those of the mIgA, dIgA, and sIgA forms for each antibody
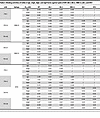 Table 3
Table 3Binding activities of mAbs in IgG, mIgA, dIgA, and sIgA forms against spike of WT, BA.1, BA.5, XBB.1.5, JN.1, and KP.3
Intranasal booster drives IgA class switching and clonal expansion of spike-specific memory B cells primed by intramuscular vaccination. We used deep sequencing to obtain BCR repertoires from PBMCs collected from the same donor before and after each vaccination, for a total of 6 time points. The BCR repertoires at each time point contained between 1.32 and 1.74 million IgH sequences. After quality control processing, an average of 30,000 unique IgH sequences were obtained per time point (Figure 3A). Next, we searched for antibodies that expressed the same clonotypes as those mucosal-derived mAbs. These clonotypes contained the same V and J genes, as well as at least 75% HCDR3 sequence identity. Before any vaccination, no clonotypes of these mAbs could be found in the BCR repertoire. After the first or second intramuscular vaccination, a low number of 719-3, 719-4, and 719-19 clonotypes were found in IgM and IgG isotypes but not in IgA isotype (Figure 3, B and C). Notably, 2 weeks after the first intranasal booster, a substantial number of 719-3, 719-4, and 719-19 in IgA clonotypes appeared, indicating marked IgA class-switching. These IgA clonotypes had higher rates of somatic hypermutation (SHM) than the IgG isotype (Figure 3C). Three months after the intranasal booster, a low number of 719-3 and 719-4 clonotypes in the IgA isotype, but not the IgG isotype, were found (Figure 3, B and C). These results suggest that the spike-specific memory B cells generated by intramuscular priming underwent IgA class-switching and clonal expansion following intranasal booster. The 719-43 clonotype was only found in the IgA isotype after intramuscular vaccination, and it increased after the intranasal booster, suggesting that IgA+ memory B cells may also undergo clonal expansion after intranasal booster (Figure 3B). The 719-43 IgA clonotype exhibited a relatively high SHM rate of 17% ± 1.5% (Figure 3C). The clonotypes 719-16, 719-25, and 719-40 could only be found as IgA after the intranasal booster. This suggests that the number of IgA+ B cells may have been increased, while the number of IgG+ B cells may have been low or largely converted to IgA+ B cells (Figure 3, B and C). The 719-4 clonotypes were the most abundant and had all 3 isotypes (IgM, IgG, and IgA) present in their BCR repertoires. This enabled us to calculate the frequency of class-switch recombination (CSR) events (Figure 3D). After intramuscular priming, the probability of CSR from IgM to IgG1 was 60%, and no CSR from IgM to IgA1 or IgG1 to IgA1 was observed. In contrast, after the intranasal booster, the probability of CSR from IgM to IgA1 increased to 75.4%, and the probability of CSR from IgG1 to IgA1 increased to 70.8% (Figure 3D).
 Figure 3
Figure 3Clonotypes of nasal mucosal mAbs in BCR repertoires at different time points after intramuscular and intranasal vaccination. (A) Sample collection scheme for sequencing BCR IgH repertoires. The schematic was drawn using BioRender. Five million peripheral mononuclear cells (PBMC) were obtained from a donor at each time point and subjected to bulk BCR sequencing as described in the methods. These time points are: before vaccination (before Vx), 3 weeks after intramuscular injection with inactivated whole-virus vaccine, 2 weeks after second intramuscular injection with inactivated whole-virus vaccine, 2 weeks after the first intranasal vaccination with Ad5-spike-BA.1, 2 weeks after the second intranasal booster, and 3 months after second intranasal booster. The total number of reads and unique reads in the heavy-chain repertoires are shown in the table. (B) The presence of 719-3, 719-4, 719-16, 719-19, 719-25, 719-40, and 719-43 clonotypes in circulating B cells at different time points after vaccination. Different shapes represent distinct clonotypes. (C) The isotype percentages and somatic hypermutation rates of 719-3, 719-4, 719-16, 719-19, 719-25, 719-40, and 719-43 clonotypes in circulating B cells at different time points after vaccination. (D) Class switch recombination (CSR) patterns of 719-4 clonotypes after intramuscular vaccination and intranasal vaccination. The color of each circle represents an Ig isotype, and the size of the circle reflects the relative number of unique Ig sequences. The arrow represents the direction of class-switching. The arrow’s thickness represents the probability of CSR, and the dashed line indicates no CSR occurrence.
We constructed a genealogical tree for clonotypes 719-3 and 719-4, as they were identified in the BCR repertoires after intramuscular vaccination and intranasal booster. The 719-3 clonotype is derived from the IGHV4-59 germline, and its B cells were generated following intramuscular vaccination at a low SHM rate of approximately 1.3%. Following the intranasal booster, the 719-3 clonotype underwent clonal expansion and IgA class-switching, and SHM rate increased to 16.7% (Supplemental Figure 4A). The 719-4 clonotype is derived from the IGHV5-51 germline, and its B cells were generated after intramuscular vaccination with an SHM rate of 1%. After the intranasal booster, the 719-4 clonotype underwent clonal expansion and IgA class switching, and SHM rate increased to 5.9% (Supplemental Figure 4B).
Spike-specific IgA+ B cells express upregulated mucosal homing receptors after intranasal booster. The tissue-specific homing of lymphocytes depends on the expression patterns of their homing receptors. It remains unclear how the intranasal booster induces localized mucosal sIgA production by modulating the homing properties of peripheral lymphocytes. To understand the phenotypic characteristics of B cells after the intranasal booster, we performed scRNA-seq on CD27+ B cells isolated from PBMCs of the same donor on days 10 and 30 following the first intranasal booster (Figure 4A and Supplemental Table 4). On day 10, four distinct B cell subtypes or states were identified, including memory B cells, plasmablasts, naive B cells, and exhausted B cells. By contrast, plasmablasts were nearly undetectable by day 30 (Figure 4B). Plasmablasts represent an antigen-induced transitional state of activated B cells. On day 10, 56.2% of plasmablasts expressed IgA, 11.7% expressed IgG, and 13.2% expressed IgM, indicating that intranasal booster predominantly promoted IgA+ B cell responses (Figure 4C). In contrast, 28.2% of memory B cells expressed IgA, 24.9% expressed IgG, and 46.3% expressed IgM by day 30 (Figure 4C). On day 10, plasmablasts express high levels of integrin α4 (ITGA4) and integrin β1 (ITGB1), which form the α4β1 integrin that can bind to vascular cell adhesion molecule-1 (VCAM-1) on mucosal epithelial cells (Figure 4, D and G). These plasmablasts also express high levels of the chemokine receptor 10 (CCR10), which is critical for homing to the nasal mucosa (Figure 4, E and H). J chain expression was the highest in plasmablasts (Figure 4, F and I). In total, 69.4% of ITGA4+ITGB1+ plasmablasts and 73.1% of CCR10+ plasmablasts express IgA, indicating IgA+ plasmablasts have upregulated expression of mucosal homing molecules (Figure 4, J and K, and Table 4). Searching for the same clonotypes as the mucosal-derived mAbs in scRNA-seq revealed 7 B cell clonotypes in the IgA isotype: 719-3, 719-4, 719-17, 719-30, 719-33, 719-39, and 719-43. Fewer B cell clonotypes (719-3, 719-4, 719-11, and 719-45) in IgG isotype could be found, and the numbers were also much less than those of the IgA isotypes (Figure 4L). These results demonstrated that the IgA class-switching, clonal expansion, and homing receptor upregulation of antigen-specific B cells occurred after the intranasal booster.
 Figure 4
Figure 4ScRNA-seq of circulating B cells on day 10 and day 30 after intranasal booster. (A and B) The t-SNE plot shows the cell clustering of 16,923 immune cells at 2 time points (A) and specific B cell types (B). The time points of cell clustering are separated according to color, with red representing day 10 and blue representing day 30 (A). Red represents plasmablasts, green represents memory B cells, gray represents naive B cells, and yellow represents exhausted B cells (B). (C) Pie charts depicting the relative proportions of IgA+ (red), IgG+ (blue), and IgM+ (gray) cells in plasmablast and memory B cell populations at day 10 and day 30 post-immunization. (D–F) The t-SNE plots show expression levels of genes ITGA4 (left panel) and ITGB1 (right panel) (D), CCR10 (E), and J CHAIN (F) in B cells on days 10 and 30 after intranasal vaccination. Color intensity represents the level of expression. (G–I) Violin plots showing transcription expression levels of α4β1 (G), CCR10 (H), and J CHAIN (I) in specific B cell types on day 10. Statistical significance was determined using the wilcoxon test, ****P < 0.0001. (J and K) Pie charts depicting the relative proportions of IgA+ (red), IgG+ (blue), and IgM+ (gray) cells in α4β1+ (J) and CCR10+ (K) plasmablast populations on day 10. (L) B cells expressing the same clonotypes as the 45 nasal mucosal mAbs in day 10 and day 30 single B cell sequencing data. Clonotypes 719-3, -4, -17, -30, -33, -39, and -43 were present in the IgA isotype (left panel), while clonotypes 719-3, -4, -11, and -45 were present in the IgG isotype (right panel).
 Table 4
Table 4Gene expression profile of plasmablasts and memory B cells on day 10 and day 30 after intranasal booster
Intranasal booster induces a transient elevation of cytokines and chemokines that facilitate B cell recruitment and antibody production in the nasal mucosa. To understand if intranasal booster induces changes in cytokines and chemokines in the nasal mucosa, which may provide insights into the induction of antigen-specific sIgA in the nasal mucosa, we collected nasal swabs from 8 volunteers on days 0, 1, 5, and 14 after the intranasal booster (Supplemental Table 6). A panel of 15 cytokines and chemokines associated with immune cell recruitment and homing was measured (Figure 5). There was a significant transient increase of C-C motif chemokine ligand 27 (CCL27), CCL28, IFN-γ, IL-2, IL-5, IL-10, IL-21, C-X-C motif chemokine ligand 11 (CXCL11), B cell activating factor (BAFF), C-X3-C motif chemokine ligand 1 (CX3CL1), and CCL22. CCL27 and CCL28 bind to CCR10 on B cells and promote the recruitment of IgA-secreting cells to the nasal mucosa (16). IFN-γ is associated with the induction of chemokines, such as CXCL11, which can promote Th1 cell recruitment (15, 25). IL-21 synergizes with TGF-β1 to promote class switch recombination (CSR) toward IgA (26, 27). IL-2, IL-5, and IL-10 promote the expansion of IgA class-switched B cells and their differentiation into IgA-secreting cells (27, 28). BAFF can promote B cell survival and antibody production (29). CCL22 promotes affinity maturation of B cells by facilitating positive selection in germinal centers (30).
 Figure 5
Figure 5Cytokine and chemokine responses in the nasal passage after intranasal booster. (A–O) Nasal swab samples were collected from 8 donors before intranasal booster, and on days 1, 5, and 14 after intranasal vaccination. The expression levels of 15 cytokines and chemokines were measured using a Luminex multiplex assay. The data are expressed as fold changes relative to prevaccination cytokine and chemokine levels. Significance was determined by wilcoxon matched-pairs signed rank test, *P < 0.05; **P < 0.01.











