Research ArticleGastroenterologyHematologyMetabolism
Open Access | ![]() 10.1172/jci.insight.184742
10.1172/jci.insight.184742
Development of rat and mouse models of heme-iron absorption
Jennifer K. Lee,1 Yue He,1 Shireen R.L. Flores,1 Regina R. Woloshun,1 Xiaoyu Wang,1 Jacob S. Shine,1 Pearl O. Ebea-Ugwuanyi,1 Sitara Sriram,1 Melissa Fraga,1 Sean Zhu,1 Yang Yu,1 Iqbal Hamza,2,3 and James F. Collins1
1Food Science & Human Nutrition Department, University of Florida, Gainesville, Florida, USA.
2Center for Blood Oxygen Transport & Hemostasis, School of Medicine, University of Maryland, Baltimore, Baltimore, Maryland, USA.
3Department of Animal and Avian Sciences, University of Maryland, College Park, Maryland, USA.
Address correspondence to: James F. Collins, Food Science & Human Nutrition Department, FSHN Bldg, #441, 572 Newell Dr., University of Florida, Gainesville, Florida, 32611, USA. Phone: 352.294.3749; Email: jfcollins@ufl.edu.
Authorship note: JKL and YH contributed equally to this work.
Find articles by Lee, J. in: PubMed | Google Scholar
1Food Science & Human Nutrition Department, University of Florida, Gainesville, Florida, USA.
2Center for Blood Oxygen Transport & Hemostasis, School of Medicine, University of Maryland, Baltimore, Baltimore, Maryland, USA.
3Department of Animal and Avian Sciences, University of Maryland, College Park, Maryland, USA.
Address correspondence to: James F. Collins, Food Science & Human Nutrition Department, FSHN Bldg, #441, 572 Newell Dr., University of Florida, Gainesville, Florida, 32611, USA. Phone: 352.294.3749; Email: jfcollins@ufl.edu.
Authorship note: JKL and YH contributed equally to this work.
Find articles by He, Y. in: PubMed | Google Scholar
1Food Science & Human Nutrition Department, University of Florida, Gainesville, Florida, USA.
2Center for Blood Oxygen Transport & Hemostasis, School of Medicine, University of Maryland, Baltimore, Baltimore, Maryland, USA.
3Department of Animal and Avian Sciences, University of Maryland, College Park, Maryland, USA.
Address correspondence to: James F. Collins, Food Science & Human Nutrition Department, FSHN Bldg, #441, 572 Newell Dr., University of Florida, Gainesville, Florida, 32611, USA. Phone: 352.294.3749; Email: jfcollins@ufl.edu.
Authorship note: JKL and YH contributed equally to this work.
Find articles by Flores, S. in: PubMed | Google Scholar
1Food Science & Human Nutrition Department, University of Florida, Gainesville, Florida, USA.
2Center for Blood Oxygen Transport & Hemostasis, School of Medicine, University of Maryland, Baltimore, Baltimore, Maryland, USA.
3Department of Animal and Avian Sciences, University of Maryland, College Park, Maryland, USA.
Address correspondence to: James F. Collins, Food Science & Human Nutrition Department, FSHN Bldg, #441, 572 Newell Dr., University of Florida, Gainesville, Florida, 32611, USA. Phone: 352.294.3749; Email: jfcollins@ufl.edu.
Authorship note: JKL and YH contributed equally to this work.
Find articles by Woloshun, R. in: PubMed | Google Scholar
1Food Science & Human Nutrition Department, University of Florida, Gainesville, Florida, USA.
2Center for Blood Oxygen Transport & Hemostasis, School of Medicine, University of Maryland, Baltimore, Baltimore, Maryland, USA.
3Department of Animal and Avian Sciences, University of Maryland, College Park, Maryland, USA.
Address correspondence to: James F. Collins, Food Science & Human Nutrition Department, FSHN Bldg, #441, 572 Newell Dr., University of Florida, Gainesville, Florida, 32611, USA. Phone: 352.294.3749; Email: jfcollins@ufl.edu.
Authorship note: JKL and YH contributed equally to this work.
Find articles by Wang, X. in: PubMed | Google Scholar
1Food Science & Human Nutrition Department, University of Florida, Gainesville, Florida, USA.
2Center for Blood Oxygen Transport & Hemostasis, School of Medicine, University of Maryland, Baltimore, Baltimore, Maryland, USA.
3Department of Animal and Avian Sciences, University of Maryland, College Park, Maryland, USA.
Address correspondence to: James F. Collins, Food Science & Human Nutrition Department, FSHN Bldg, #441, 572 Newell Dr., University of Florida, Gainesville, Florida, 32611, USA. Phone: 352.294.3749; Email: jfcollins@ufl.edu.
Authorship note: JKL and YH contributed equally to this work.
Find articles by Shine, J. in: PubMed | Google Scholar
1Food Science & Human Nutrition Department, University of Florida, Gainesville, Florida, USA.
2Center for Blood Oxygen Transport & Hemostasis, School of Medicine, University of Maryland, Baltimore, Baltimore, Maryland, USA.
3Department of Animal and Avian Sciences, University of Maryland, College Park, Maryland, USA.
Address correspondence to: James F. Collins, Food Science & Human Nutrition Department, FSHN Bldg, #441, 572 Newell Dr., University of Florida, Gainesville, Florida, 32611, USA. Phone: 352.294.3749; Email: jfcollins@ufl.edu.
Authorship note: JKL and YH contributed equally to this work.
Find articles by Ebea-Ugwuanyi, P. in: PubMed | Google Scholar
1Food Science & Human Nutrition Department, University of Florida, Gainesville, Florida, USA.
2Center for Blood Oxygen Transport & Hemostasis, School of Medicine, University of Maryland, Baltimore, Baltimore, Maryland, USA.
3Department of Animal and Avian Sciences, University of Maryland, College Park, Maryland, USA.
Address correspondence to: James F. Collins, Food Science & Human Nutrition Department, FSHN Bldg, #441, 572 Newell Dr., University of Florida, Gainesville, Florida, 32611, USA. Phone: 352.294.3749; Email: jfcollins@ufl.edu.
Authorship note: JKL and YH contributed equally to this work.
Find articles by Sriram, S. in: PubMed | Google Scholar
1Food Science & Human Nutrition Department, University of Florida, Gainesville, Florida, USA.
2Center for Blood Oxygen Transport & Hemostasis, School of Medicine, University of Maryland, Baltimore, Baltimore, Maryland, USA.
3Department of Animal and Avian Sciences, University of Maryland, College Park, Maryland, USA.
Address correspondence to: James F. Collins, Food Science & Human Nutrition Department, FSHN Bldg, #441, 572 Newell Dr., University of Florida, Gainesville, Florida, 32611, USA. Phone: 352.294.3749; Email: jfcollins@ufl.edu.
Authorship note: JKL and YH contributed equally to this work.
Find articles by Fraga, M. in: PubMed | Google Scholar
1Food Science & Human Nutrition Department, University of Florida, Gainesville, Florida, USA.
2Center for Blood Oxygen Transport & Hemostasis, School of Medicine, University of Maryland, Baltimore, Baltimore, Maryland, USA.
3Department of Animal and Avian Sciences, University of Maryland, College Park, Maryland, USA.
Address correspondence to: James F. Collins, Food Science & Human Nutrition Department, FSHN Bldg, #441, 572 Newell Dr., University of Florida, Gainesville, Florida, 32611, USA. Phone: 352.294.3749; Email: jfcollins@ufl.edu.
Authorship note: JKL and YH contributed equally to this work.
Find articles by Zhu, S. in: PubMed | Google Scholar
1Food Science & Human Nutrition Department, University of Florida, Gainesville, Florida, USA.
2Center for Blood Oxygen Transport & Hemostasis, School of Medicine, University of Maryland, Baltimore, Baltimore, Maryland, USA.
3Department of Animal and Avian Sciences, University of Maryland, College Park, Maryland, USA.
Address correspondence to: James F. Collins, Food Science & Human Nutrition Department, FSHN Bldg, #441, 572 Newell Dr., University of Florida, Gainesville, Florida, 32611, USA. Phone: 352.294.3749; Email: jfcollins@ufl.edu.
Authorship note: JKL and YH contributed equally to this work.
Find articles by Yu, Y. in: PubMed | Google Scholar
1Food Science & Human Nutrition Department, University of Florida, Gainesville, Florida, USA.
2Center for Blood Oxygen Transport & Hemostasis, School of Medicine, University of Maryland, Baltimore, Baltimore, Maryland, USA.
3Department of Animal and Avian Sciences, University of Maryland, College Park, Maryland, USA.
Address correspondence to: James F. Collins, Food Science & Human Nutrition Department, FSHN Bldg, #441, 572 Newell Dr., University of Florida, Gainesville, Florida, 32611, USA. Phone: 352.294.3749; Email: jfcollins@ufl.edu.
Authorship note: JKL and YH contributed equally to this work.
Find articles by Hamza, I. in: PubMed | Google Scholar
1Food Science & Human Nutrition Department, University of Florida, Gainesville, Florida, USA.
2Center for Blood Oxygen Transport & Hemostasis, School of Medicine, University of Maryland, Baltimore, Baltimore, Maryland, USA.
3Department of Animal and Avian Sciences, University of Maryland, College Park, Maryland, USA.
Address correspondence to: James F. Collins, Food Science & Human Nutrition Department, FSHN Bldg, #441, 572 Newell Dr., University of Florida, Gainesville, Florida, 32611, USA. Phone: 352.294.3749; Email: jfcollins@ufl.edu.
Authorship note: JKL and YH contributed equally to this work.
Find articles by Collins, J. in: PubMed | Google Scholar
Published June 9, 2025 - More info
JCI Insight. 2025;10(11):e184742. https://doi.org/10.1172/jci.insight.184742.
© 2025 Lee et al. This work is licensed under the Creative Commons Attribution 4.0 International License. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
Received: July 11, 2024; Accepted: April 23, 2025

-
Results
Iron depletion-repletion studies in rats. In rat pilot study 1, the ability of dietary HI to mitigate IDA was assessed in 3 common strains of laboratory rats (Rattus norvegicus): Wistar, Lewis, and S-D. Wistar and S-D rats are outbred, while Lewis rats are inbred. Results revealed that the S-D rat exhibited the most significant increase in blood Hb levels upon short-term repletion with an HI-enriched diet (Supplemental Figure 1; supplemental material available online with this article; https://doi.org/10.1172/jci.insight.184742DS1), so this strain was selected for further experimentation. In rat pilot study 2, a longer-term experiment was undertaken. Results demonstrated that repletion of iron-deficient, anemic S-D rats with a 50-ppm NHI diet or an HI-enriched diet with 50 ppm total iron both effectively resolved IDA. Although the Hb recovery rate was apparently slower in rats consuming the HI diet, Hb levels were normalized in both dietary groups by the end of the 26-day repletion period (Figure 1A). Both diets also restored serum (transport) iron and liver (storage) iron and transferrin saturation (TSAT) to control levels. Spleen iron, however, was not fully replenished in the HI diet–fed rats (Figure 1B), suggesting that the animals remained moderately iron deficient. These experiments provided the rationale for further testing and development of the S-D rat as a model of HI absorption/utilization.
 Figure 1
Figure 1Rat pilot study 2: An HI diet was effective at correcting the anemia in iron-depleted, S-D rats. Weanling, female rats were fed a low-iron diet for 2 weeks (~3 ppm NHI) and then repleted (R) with a 7-ppm NHI (internal control) diet, a 50-ppm NHI (control) diet, or a 50-ppm HI–enriched (experimental) diet. Additional control groups were fed the iron-adequate (50 ppm NHI) diet or the low-iron diet (3 ppm NHI) throughout. Rats were bled every 4 days and sacrificed 26 days after initiation of the iron-repletion phase. Shown are Hb levels at each time point (A) and tissue/serum NHI levels and TSAT at the end of the study (B). Data are mean ± SD for n = 4–5 rats per group and were analyzed by 2-way (A) or 1-way (B) ANOVA followed by Tukey’s multiple-comparison test. Groups with different letters vary significantly. Significant 2-way interaction (A) and main effect (B) P values are indicated in each panel. A: BL, baseline, prior to iron deprivation; 0, after 2 weeks of low-iron feeding; 2 days, 6 days, etc., indicates repletion period in days.
Next, since the 50-ppm HI diet did not fully correct the iron deficiency in iron-depleted rats, a subsequent experiment utilized HI-enriched diets containing total iron up to 100 ppm, which is still within the upper range of physiological intakes but well below the iron content of most commercial rodent chows (which may contain ≥300 ppm Fe). Outcomes of this experiment showed that a 50-ppm NHI (control) repletion diet, and 75- and 100-ppm HI–enriched (experimental) repletion diets, all corrected the anemia within 8 days (Figure 2A); however, the 50-ppm HI and 15-ppm NHI repletion diets did not. The 50-ppm NHI repletion diet, and the 75- and 100-ppm HI repletion diets, were also equally effective at restoring hematocrit (Hct), TSAT, serum, liver, spleen, bone marrow, kidney, and heart iron levels (Figure 2, B–I), as well as serum hepcidin and liver hepcidin, renal erythropoietin (Epo), and bone marrow erythroferrone (Erfe) mRNA expression (Figure 3). ERFE is hormone that is secreted by developing erythrocytes in the bone marrow, under the influence of EPO, that downregulates Hamp gene transcription in hepatocytes, thus reducing circulating hepcidin levels (39).
 Figure 2
Figure 2The 50-ppm NHI and 75-ppm HI repletion diets were equally effective at correcting IDA in S-D rats. Weanling, female rats were fed a low-iron diet (3 ppm NHI) for 2 weeks and then repleted (R) with 50-, 75-, or 100-ppm HI–enriched experimental diets, or a 50-ppm NHI (iron adequate) control diet. The 15-ppm NHI (internal control) diet matched the NHI content of the 100-ppm HI–enriched diet (i.e., 85 ppm HI and 15 ppm NHI). Additional control groups were fed the iron-adequate or the low-iron diet throughout. Rats were bled every 4 days and sacrificed 8 days after initiation of the iron-repletion phase. Hb levels are shown at each time point (A). Also shown are Hct (B), serum TSAT (C), and serum (D), liver (E), spleen (F), bone marrow (G), kidney (H) and heart (I) NHI levels in experimental rats at the termination of the experiment. Results are presented as a line graph (A) or box-and-whisker plots (B–I) for n = 3–6 rats per group. Data were analyzed by 2-way (A) or 1-way (B–I) ANOVA followed by Tukey’s multiple-comparison test. Groups with different letters vary significantly. Significant main effect and 2-way interaction P values are shown in each panel. A: BL, baseline, prior to iron deprivation; 0, after 2 weeks of low-iron feeding; 4d and 8 d indicate repletion period in days.
 Figure 3
Figure 3A 75-ppm HI diet was effective at normalizing serum hepcidin and EPO levels in iron-deprived S-D rats. Experimental details are the same as for those described in the Figure 2 legend. Shown are serum hepcidin (A), liver hepcidin mRNA expression (B), serum EPO (C), kidney Epo mRNA expression (D), and bone marrow Erfe mRNA expression (E). Results are presented as box-and-whisker plots for n = 3–6 rats per group. Data were analyzed by 1-way ANOVA followed by Tukey’s multiple-comparison test. Groups with different letters vary significantly. Significant main effect P values are shown in each panel. (R), repletion group.
Collectively, these experiments revealed that a 75-ppm HI–enriched diet, containing approximately 64 ppm HI and 11 ppm NHI, provided sufficient iron to fully mitigate severe IDA in adolescent S-D rats. Since the internal control (15 ppm) NHI diet was ineffective at correcting the IDA in these rats, we concluded that S-D rats efficiently assimilated and utilized iron derived from heme.
Iron depletion-repletion studies in mice. Encouraged by outcomes in S-D rats, we next assessed HI assimilation by iron-depleted B6 mice, a well-established mouse strain for iron metabolism studies. The rationale for studying mice was 2-fold: (a) HI absorption has been described as an inefficient process in mice (28), based on scant experimental evidence, and this is now the generalized perception in the field. As such, no extensively characterized mouse models of HI absorption have been developed to date; and (b) a multitude of genetically engineered mouse strains are commercially available, which could accelerate future discovery. We first sought to define the optimal dietary iron concentration to use for mouse experiments. In mouse pilot study 1, experimental HI-enriched diets containing 50–100 ppm total iron were utilized, since we postulated that dietary iron requirements would be similar for rats and mice. However, outcomes demonstrated that all of the experimental HI-enriched diets, and the control 50-ppm NHI diet, rapidly corrected IDA in anemic B6 mice (Hb ~11 g/dL) within 8 days (Supplemental Figure 2). This suggested that dietary iron should be lowered for experiments in mice, so the HI-enriched diets were reformulated with total iron ranging from 15 to 30 ppm. These diets were fed to iron-deficient, anemic mice (Hb ~5 g/dL) in mouse pilot study 2. Notably, this level of anemia better reflected the severe anemia in the rat iron depletion/repletion experiments described above. Results of this experiment demonstrated that 30 ppm HI was insufficient to fully restore Hb levels (Supplemental Figure 3), so diets were reformulated again with total iron up to 60 ppm. These dietary iron levels turned out to be appropriate, as the 60-ppm HI repletion diet increased blood Hb in iron-deprived, anemic mice (Hb ~5 g/dL) at essentially the same rate as the control 50-ppm NHI repletion diet (Figure 4A). The 15- and 30-ppm HI diets, and the 9-ppm NHI internal control diet, however, did not correct the anemia, even after 24 days of repletion. The 60-ppm HI–enriched diet also normalized Hct, TSAT, and serum and spleen iron levels, just as effectively as the iron-adequate, 50-ppm NHI diet (Figure 4, B–D, and F). None of the experimental HI diets (including the 100-ppm HI diet used in mouse pilot study 1), however, were able to fully replenish liver iron stores (Figure 4E). The 60-ppm HI diet also normalized liver Hamp and bone marrow Erfe mRNA expression (Figure 4, G and I). Renal Epo mRNA expression did not vary significantly between groups (Figure 4H). B6 mice thus effectively utilized dietary HI to recover from severe IDA.
 Figure 4
Figure 4A 60-ppm HI diet was effective at correcting the IDA in iron-depleted C57BL/6 mice. Female weanling mice were fed a low-iron diet (3 ppm NHI) for 3 weeks and then switched to control repletion diets (50 ppm and 9 ppm NHI), or 1 of 3 different HI-enriched (experimental) repletion diets (15, 30, or 60 ppm iron). Additional control groups consumed an iron-adequate diet (50 ppm NHI) or the low-iron diet throughout. Mice were bled every 4 days and sacrificed 24 days after initiation of the iron-repletion phase. Hb levels are shown at each time point (A). Also shown are Hct (B), serum TSAT (C), and serum (D), liver (E) and spleen (F) NHI from mice at the termination of the experiment. Also shown are liver Hamp (G), kidney Epo (H), and bone marrow Erfe mRNA expression (I). Results are presented as a line graph (A) or box-and-whisker plots (B–I) for n = 5–6 mice per group. Data were analyzed by 2-way (A) or 1-way (B–I) ANOVA followed by Tukey’s multiple-comparison test. Groups with different letters vary significantly. Significant 2-way interaction (A) and main effect (B–I) P values are shown in each panel. A: BL, baseline, prior to iron deprivation; 0, after 3 weeks of low-iron feeding; 4d, 8d, etc., indicate repletion (R) period in days.
HI utilization during pregnancy by S-D rats and B6 mice. Extra dietary iron is required to support pregnancy, lactation, and neonatal development in humans and rodents. To investigate whether dietary heme could provide this extra iron, we utilized an experimental 50-ppm HI–enriched diet, containing approximately 43 ppm HI and 7 ppm NHI, and an iron-adequate, 50-ppm NHI control diet. Importantly, the minimal amount of NHI iron recommended for rats and mice in the AIN-93G diet is 35 ppm (40, 41), which is formulated for the growth, pregnancy, and lactation phases of development. The 7-ppm background NHI in the 50-ppm HI–enriched diet is thus well below this minimum requirement, so inclusion of a 7-ppm NHI internal control diet was unnecessary. Female S-D rats were fed these diets for 1 week prior to mating and conception, and throughout pregnancy and lactation. Outcomes showed that many parameters were invariable between dietary groups, including the number and sex distribution of pups, blood Hb levels and TSAT in dams and pups, as well as serum, liver, and spleen NHI in pups (Tables 1 and 2). Some differences were also noted between groups, including lower body weights in dams (~20%) and pups (20%–25%) in the HI groups. Dams consuming the HI diet also had lower serum iron (by ~40%), but surprisingly, liver (by ~3-fold) and spleen (by ~2-fold) iron were elevated significantly, as compared with dams fed the NHI diet. Tissue distribution of iron thus varied by dietary source (HI vs. NHI), consistent with findings from a recent publication (31). Increasing total iron up to 75 ppm may have prevented the noted growth impairment in dams and pups, since this amount of dietary iron was the minimal amount requirement to fully support recovery from severe IDA in S-D rats (Figures 3 and 4).
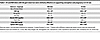 Table 1
Table 1HI and NHI diets with 50 ppm total iron were similarly effective at supporting conception and pregnancy in S-D rats
 Table 2
Table 2HI and NHI diets with 50 ppm total iron were similarly effective at supporting pup development in S-D rats
An identical pregnancy study in B6 mice showed that there were no noted differences in any measured parameters in dams consuming either diet (Table 3). In the pups, almost all outcomes were also invariable between dietary groups, with the only difference being a 13% (possibly unimportant) reduction in Hb levels in males (Table 4). The 50-ppm HI diet thus fully supported normal fertility, pregnancy, lactation, and development of neonates in mice. Collectively, these data demonstrated that S-D rats and B6 mice efficiently utilized iron derived from dietary Hb/heme to meet the enhanced iron requirements necessary to support successful pregnancy outcomes.
 Table 3
Table 3HI and NHI diets with 50 ppm total iron were equally effective at supporting conception and pregnancy in C57BL/6 mice
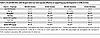 Table 4
Table 4HI and NHI diets with 50 ppm total iron were equally effective at supporting pup development in C57BL/6 mice
Defining iron requirements for Hamp-KO B6 mice and S-D rats, and iron loading study. HH is a heritable, iron-loading disorder in humans, most commonly caused by mutations in the HFE gene (42). Mutations in HAMP, encoding hepcidin, underlie a rare, severe early-onset form of the disease (43, 44). In HH, iron overload ensues due to low hepcidin, which results in inappropriately elevated intestinal iron absorption and pathological iron accumulation in parenchymal tissues. Evidence in the scientific literature supports the concept that dietary HI and NHI are both hyperabsorbed in patients with HH (45, 46). Hamp-KO mice are an excellent model of HH (47). Since our studies revealed efficient HI absorption and utilization by rats and mice, we next examined whether dietary HI could also precipitate iron loading in Hamp-KO S-D rats (48) and B6 mice. However, it was first necessary to define the dietary iron requirements for Hamp-KO rats and mice, since they hyperabsorb dietary iron.
Weanling, WT and Hamp-KO mice were fed diets with variable NHI content (5–50 ppm) for 6 weeks (Figure 5A). Consumption of the diets with 30 and 50 ppm total iron caused iron loading in Hamp KOs, as indicated by elevated serum ferritin, a biomarker for body iron stores (49), and increased liver iron content, as compared with WT mice consuming the same diets. Hamp-KO mice consuming the 5- and 15-ppm NHI diets were, however, protected from iron overload (Figure 5, B–F). Given these outcomes, a subsequent iron loading study was undertaken using NHI (control) and HI-enriched (experimental) diets with 50 ppm total iron. The use of an internal control diet with 7 ppm NHI, to match the NHI content of the HI-enriched diet, was deemed unnecessary. Outcomes from a subsequent 6-week, iron-loading study showed that Hamp-KO mice consuming both diets had elevated TSAT, and higher serum and tissue NHI levels, indicative of iron overload (Figure 6). In the liver, pancreas, and heart, iron loading was more pronounced in the NHI diet group, and iron loading in the kidney was only seen in this group. Nonetheless, these data demonstrated that NHI and HI both contribute to iron loading in Hamp-KO mice, consistent with observations made in humans (45, 46). These data further supported the concept that, under these experimental conditions, B6 mice assimilated iron from dietary heme.
 Figure 5
Figure 5Consumption of a 15-ppm NHI diet prevents iron overload in Hamp-KO C57BL/6 mice. Male (blue symbols) and female (red symbols) WT (circles) and Hamp-KO (squares) mice were fed diets with variable NHI content for 6 weeks after weaning. Shown are experimental design (A), serum NHI levels (B), serum TSAT (C), serum ferritin (D), and spleen (E) and liver (F) NHI content. Data are presented as box-and-whisker plots for n = 6 mice/group and were analyzed by 2-way ANOVA followed by Tukey’s multiple-comparison test. Asterisks denote significant differences between the WT and KOs within a diet group. *P < 0.05; ***P < 0.001; ****P < 0.0001. Significant 2-way interaction and main effect P values are shown in each panel.
 Figure 6
Figure 6Consumption of a 50-ppm NHI diet or a 50-ppm HI–enriched diet leads to iron loading in Hamp-KO C57BL/6 mice. Female WT and Hamp-KO mice were weaned onto a control diet with 50 ppm NHI (blue circles), or an experimental 50-ppm HI–enriched diet (red squares), and then sacrificed 6 weeks later. Shown are serum TSAT (A), and serum (B), liver (C), pancreas (D), kidney (E), and heart (F) NHI levels, which were determined in experimental mice at the termination of the experiment. Data were analyzed by 2-way ANOVA followed by Tukey’s multiple-comparison test. Results are presented as box-and-whisker plots for n = 6 mice per group. Groups with different letters vary significantly. Significant 2-way interaction and main effect P values are shown in each panel.
Parallel experiments in Hamp-KO S-D rats showed similar outcomes. As in mice, consumption of the 50- and 30-ppm NHI diets led to increases in serum and liver iron content, and TSAT, indicative of iron loading in the KOs, but rats consuming the 5- and 15-ppm NHI diets did not load excess iron (Figure 7). As in mice, iron overload was more pronounced when rats consumed the 50-ppm iron diet. A subsequent iron loading study showed that both the NHI and HI-enriched diets with 50 ppm total iron increased serum and liver iron levels in Hamp KOs (Figure 8). Changes in pancreatic and heart iron levels, although statistically significant, were minor and less likely to be biologically relevant. Renal iron content was lower in both KO groups, but no differences were noted when comparing diets within each genotype. NHI and HI thus both contributed to the iron loading caused by excessive intestinal iron absorption in Hamp-KO S-D rats and B6 mice.
 Figure 7
Figure 7Consumption of a 15-ppm NHI diet prevents iron overload in Hamp-KO S-D rats. WT and KO rats of both sexes were fed diets with variable NHI content for 6 weeks after weaning and then sacrificed for analysis. Shown are serum NHI levels (A), TSAT (B), serum ferritin (C), and spleen (D), liver (E) and bone marrow (F) NHI content. Data are presented as box-and-whisker plots for n = 5–6 rats/group and were analyzed by 2-way ANOVA followed by Tukey’s multiple-comparison test. Asterisks denote significant differences between the WT and KOs within a diet group. *P < 0.05; **P < 0.01; ***P < 0.001; ****P < 0.0001. Significant 2-way interaction and main effect P values are shown in each panel.
 Figure 8
Figure 8Consumption of a 50-ppm NHI diet or a 50-ppm HI–enriched diet leads to iron loading in Hamp-KO S-D rats. Female WT and Hamp-KO rats were weaned onto a control diet with 50 ppm NHI (blue circles), or an experimental 50-ppm HI–enriched diet (red squares), and then sacrificed after 6 weeks on the diets. Shown are serum TSAT (A), and serum (B), liver (C), pancreas (D), kidney (E), and heart (F) NHI levels, which were determined in experimental rats at the termination of the experiment. Results are presented as box-and-whisker plots for n = 6 rats per group. Data were analyzed by 2-way ANOVA followed by Tukey’s multiple-comparison test. Groups with different letters vary significantly. Significant 2-way interaction and main effect P values are shown in each panel.