Clinical Research and Public HealthGastroenterologyMicrobiology
Open Access | ![]() 10.1172/jci.insight.188993
10.1172/jci.insight.188993
Host-microbiome determinants of ready-to-use supplemental food efficacy in acute childhood malnutrition
Zehra Jamil,1,2 Gabriel F. Hanson,3 Junaid Iqbal,4 G. Brett Moreau,5 Najeeha Talat Iqbal,1,4 Sheraz Ahmed,4 Aneeta Hotwani,4 Furqan Kabir,4 Fayaz Umrani,4 Kamran Sadiq,4 Kumail Ahmed,4 Indika Mallawaarachchi,6 Jennie Z. Ma,6 Fatima Aziz,4 S. Asad Ali,2 and Sean R. Moore7
1Department of Biological & Biomedical Sciences and
2Department of Community & Health Sciences, Aga Khan University, Karachi, Pakistan.
3Department of Biomedical Engineering, University of Virginia, Charlottesville, Virginia, USA.
4Department of Pediatrics & Child Health, Aga Khan University, Karachi, Pakistan.
5Division of Infectious Diseases and International Health, Department of Medicine, University of Virginia School of Medicine, Charlottesville, Virginia, USA.
6Department of Public Health Sciences, University of Virginia, Charlottesville, Virginia, USA.
7Division of Gastroenterology, Hepatology and Nutrition, Cincinnati Children’s Hospital, Cincinnati, Ohio, USA.
Address correspondence to: Syed Asad Ali, Chair, Department of Community Health Sciences, Professor, Pediatric Infectious Disease, Aga Khan University, National Stadium Road PO Box 3500, Karachi 74800, Pakistan. Email: asad.ali@aku.edu. Or to: Sean Ryan Moore, Division Director, Division of Gastroenterology, Hepatology and Nutrition, Professor, UC Department of Pediatrics, Cincinnati Children’s Hospital Medical Center, 3333 Burnet Ave, Cincinnati, Ohio 45229, USA. Email: sean.moore@cchmc.org.
Authorship note: ZJ and GFH contributed equally to this work.
Find articles by Jamil, Z. in: PubMed | Google Scholar
1Department of Biological & Biomedical Sciences and
2Department of Community & Health Sciences, Aga Khan University, Karachi, Pakistan.
3Department of Biomedical Engineering, University of Virginia, Charlottesville, Virginia, USA.
4Department of Pediatrics & Child Health, Aga Khan University, Karachi, Pakistan.
5Division of Infectious Diseases and International Health, Department of Medicine, University of Virginia School of Medicine, Charlottesville, Virginia, USA.
6Department of Public Health Sciences, University of Virginia, Charlottesville, Virginia, USA.
7Division of Gastroenterology, Hepatology and Nutrition, Cincinnati Children’s Hospital, Cincinnati, Ohio, USA.
Address correspondence to: Syed Asad Ali, Chair, Department of Community Health Sciences, Professor, Pediatric Infectious Disease, Aga Khan University, National Stadium Road PO Box 3500, Karachi 74800, Pakistan. Email: asad.ali@aku.edu. Or to: Sean Ryan Moore, Division Director, Division of Gastroenterology, Hepatology and Nutrition, Professor, UC Department of Pediatrics, Cincinnati Children’s Hospital Medical Center, 3333 Burnet Ave, Cincinnati, Ohio 45229, USA. Email: sean.moore@cchmc.org.
Authorship note: ZJ and GFH contributed equally to this work.
Find articles by Hanson, G. in: PubMed | Google Scholar
1Department of Biological & Biomedical Sciences and
2Department of Community & Health Sciences, Aga Khan University, Karachi, Pakistan.
3Department of Biomedical Engineering, University of Virginia, Charlottesville, Virginia, USA.
4Department of Pediatrics & Child Health, Aga Khan University, Karachi, Pakistan.
5Division of Infectious Diseases and International Health, Department of Medicine, University of Virginia School of Medicine, Charlottesville, Virginia, USA.
6Department of Public Health Sciences, University of Virginia, Charlottesville, Virginia, USA.
7Division of Gastroenterology, Hepatology and Nutrition, Cincinnati Children’s Hospital, Cincinnati, Ohio, USA.
Address correspondence to: Syed Asad Ali, Chair, Department of Community Health Sciences, Professor, Pediatric Infectious Disease, Aga Khan University, National Stadium Road PO Box 3500, Karachi 74800, Pakistan. Email: asad.ali@aku.edu. Or to: Sean Ryan Moore, Division Director, Division of Gastroenterology, Hepatology and Nutrition, Professor, UC Department of Pediatrics, Cincinnati Children’s Hospital Medical Center, 3333 Burnet Ave, Cincinnati, Ohio 45229, USA. Email: sean.moore@cchmc.org.
Authorship note: ZJ and GFH contributed equally to this work.
Find articles by Iqbal, J. in: PubMed | Google Scholar
1Department of Biological & Biomedical Sciences and
2Department of Community & Health Sciences, Aga Khan University, Karachi, Pakistan.
3Department of Biomedical Engineering, University of Virginia, Charlottesville, Virginia, USA.
4Department of Pediatrics & Child Health, Aga Khan University, Karachi, Pakistan.
5Division of Infectious Diseases and International Health, Department of Medicine, University of Virginia School of Medicine, Charlottesville, Virginia, USA.
6Department of Public Health Sciences, University of Virginia, Charlottesville, Virginia, USA.
7Division of Gastroenterology, Hepatology and Nutrition, Cincinnati Children’s Hospital, Cincinnati, Ohio, USA.
Address correspondence to: Syed Asad Ali, Chair, Department of Community Health Sciences, Professor, Pediatric Infectious Disease, Aga Khan University, National Stadium Road PO Box 3500, Karachi 74800, Pakistan. Email: asad.ali@aku.edu. Or to: Sean Ryan Moore, Division Director, Division of Gastroenterology, Hepatology and Nutrition, Professor, UC Department of Pediatrics, Cincinnati Children’s Hospital Medical Center, 3333 Burnet Ave, Cincinnati, Ohio 45229, USA. Email: sean.moore@cchmc.org.
Authorship note: ZJ and GFH contributed equally to this work.
Find articles by
Moreau, G.
in:
PubMed
|
Google Scholar
|

1Department of Biological & Biomedical Sciences and
2Department of Community & Health Sciences, Aga Khan University, Karachi, Pakistan.
3Department of Biomedical Engineering, University of Virginia, Charlottesville, Virginia, USA.
4Department of Pediatrics & Child Health, Aga Khan University, Karachi, Pakistan.
5Division of Infectious Diseases and International Health, Department of Medicine, University of Virginia School of Medicine, Charlottesville, Virginia, USA.
6Department of Public Health Sciences, University of Virginia, Charlottesville, Virginia, USA.
7Division of Gastroenterology, Hepatology and Nutrition, Cincinnati Children’s Hospital, Cincinnati, Ohio, USA.
Address correspondence to: Syed Asad Ali, Chair, Department of Community Health Sciences, Professor, Pediatric Infectious Disease, Aga Khan University, National Stadium Road PO Box 3500, Karachi 74800, Pakistan. Email: asad.ali@aku.edu. Or to: Sean Ryan Moore, Division Director, Division of Gastroenterology, Hepatology and Nutrition, Professor, UC Department of Pediatrics, Cincinnati Children’s Hospital Medical Center, 3333 Burnet Ave, Cincinnati, Ohio 45229, USA. Email: sean.moore@cchmc.org.
Authorship note: ZJ and GFH contributed equally to this work.
Find articles by Iqbal, N. in: PubMed | Google Scholar
1Department of Biological & Biomedical Sciences and
2Department of Community & Health Sciences, Aga Khan University, Karachi, Pakistan.
3Department of Biomedical Engineering, University of Virginia, Charlottesville, Virginia, USA.
4Department of Pediatrics & Child Health, Aga Khan University, Karachi, Pakistan.
5Division of Infectious Diseases and International Health, Department of Medicine, University of Virginia School of Medicine, Charlottesville, Virginia, USA.
6Department of Public Health Sciences, University of Virginia, Charlottesville, Virginia, USA.
7Division of Gastroenterology, Hepatology and Nutrition, Cincinnati Children’s Hospital, Cincinnati, Ohio, USA.
Address correspondence to: Syed Asad Ali, Chair, Department of Community Health Sciences, Professor, Pediatric Infectious Disease, Aga Khan University, National Stadium Road PO Box 3500, Karachi 74800, Pakistan. Email: asad.ali@aku.edu. Or to: Sean Ryan Moore, Division Director, Division of Gastroenterology, Hepatology and Nutrition, Professor, UC Department of Pediatrics, Cincinnati Children’s Hospital Medical Center, 3333 Burnet Ave, Cincinnati, Ohio 45229, USA. Email: sean.moore@cchmc.org.
Authorship note: ZJ and GFH contributed equally to this work.
Find articles by
Ahmed, S.
in:
PubMed
|
Google Scholar
|
1Department of Biological & Biomedical Sciences and
2Department of Community & Health Sciences, Aga Khan University, Karachi, Pakistan.
3Department of Biomedical Engineering, University of Virginia, Charlottesville, Virginia, USA.
4Department of Pediatrics & Child Health, Aga Khan University, Karachi, Pakistan.
5Division of Infectious Diseases and International Health, Department of Medicine, University of Virginia School of Medicine, Charlottesville, Virginia, USA.
6Department of Public Health Sciences, University of Virginia, Charlottesville, Virginia, USA.
7Division of Gastroenterology, Hepatology and Nutrition, Cincinnati Children’s Hospital, Cincinnati, Ohio, USA.
Address correspondence to: Syed Asad Ali, Chair, Department of Community Health Sciences, Professor, Pediatric Infectious Disease, Aga Khan University, National Stadium Road PO Box 3500, Karachi 74800, Pakistan. Email: asad.ali@aku.edu. Or to: Sean Ryan Moore, Division Director, Division of Gastroenterology, Hepatology and Nutrition, Professor, UC Department of Pediatrics, Cincinnati Children’s Hospital Medical Center, 3333 Burnet Ave, Cincinnati, Ohio 45229, USA. Email: sean.moore@cchmc.org.
Authorship note: ZJ and GFH contributed equally to this work.
Find articles by Hotwani, A. in: PubMed | Google Scholar
1Department of Biological & Biomedical Sciences and
2Department of Community & Health Sciences, Aga Khan University, Karachi, Pakistan.
3Department of Biomedical Engineering, University of Virginia, Charlottesville, Virginia, USA.
4Department of Pediatrics & Child Health, Aga Khan University, Karachi, Pakistan.
5Division of Infectious Diseases and International Health, Department of Medicine, University of Virginia School of Medicine, Charlottesville, Virginia, USA.
6Department of Public Health Sciences, University of Virginia, Charlottesville, Virginia, USA.
7Division of Gastroenterology, Hepatology and Nutrition, Cincinnati Children’s Hospital, Cincinnati, Ohio, USA.
Address correspondence to: Syed Asad Ali, Chair, Department of Community Health Sciences, Professor, Pediatric Infectious Disease, Aga Khan University, National Stadium Road PO Box 3500, Karachi 74800, Pakistan. Email: asad.ali@aku.edu. Or to: Sean Ryan Moore, Division Director, Division of Gastroenterology, Hepatology and Nutrition, Professor, UC Department of Pediatrics, Cincinnati Children’s Hospital Medical Center, 3333 Burnet Ave, Cincinnati, Ohio 45229, USA. Email: sean.moore@cchmc.org.
Authorship note: ZJ and GFH contributed equally to this work.
Find articles by Kabir, F. in: PubMed | Google Scholar
1Department of Biological & Biomedical Sciences and
2Department of Community & Health Sciences, Aga Khan University, Karachi, Pakistan.
3Department of Biomedical Engineering, University of Virginia, Charlottesville, Virginia, USA.
4Department of Pediatrics & Child Health, Aga Khan University, Karachi, Pakistan.
5Division of Infectious Diseases and International Health, Department of Medicine, University of Virginia School of Medicine, Charlottesville, Virginia, USA.
6Department of Public Health Sciences, University of Virginia, Charlottesville, Virginia, USA.
7Division of Gastroenterology, Hepatology and Nutrition, Cincinnati Children’s Hospital, Cincinnati, Ohio, USA.
Address correspondence to: Syed Asad Ali, Chair, Department of Community Health Sciences, Professor, Pediatric Infectious Disease, Aga Khan University, National Stadium Road PO Box 3500, Karachi 74800, Pakistan. Email: asad.ali@aku.edu. Or to: Sean Ryan Moore, Division Director, Division of Gastroenterology, Hepatology and Nutrition, Professor, UC Department of Pediatrics, Cincinnati Children’s Hospital Medical Center, 3333 Burnet Ave, Cincinnati, Ohio 45229, USA. Email: sean.moore@cchmc.org.
Authorship note: ZJ and GFH contributed equally to this work.
Find articles by Umrani, F. in: PubMed | Google Scholar
1Department of Biological & Biomedical Sciences and
2Department of Community & Health Sciences, Aga Khan University, Karachi, Pakistan.
3Department of Biomedical Engineering, University of Virginia, Charlottesville, Virginia, USA.
4Department of Pediatrics & Child Health, Aga Khan University, Karachi, Pakistan.
5Division of Infectious Diseases and International Health, Department of Medicine, University of Virginia School of Medicine, Charlottesville, Virginia, USA.
6Department of Public Health Sciences, University of Virginia, Charlottesville, Virginia, USA.
7Division of Gastroenterology, Hepatology and Nutrition, Cincinnati Children’s Hospital, Cincinnati, Ohio, USA.
Address correspondence to: Syed Asad Ali, Chair, Department of Community Health Sciences, Professor, Pediatric Infectious Disease, Aga Khan University, National Stadium Road PO Box 3500, Karachi 74800, Pakistan. Email: asad.ali@aku.edu. Or to: Sean Ryan Moore, Division Director, Division of Gastroenterology, Hepatology and Nutrition, Professor, UC Department of Pediatrics, Cincinnati Children’s Hospital Medical Center, 3333 Burnet Ave, Cincinnati, Ohio 45229, USA. Email: sean.moore@cchmc.org.
Authorship note: ZJ and GFH contributed equally to this work.
Find articles by Sadiq, K. in: PubMed | Google Scholar
1Department of Biological & Biomedical Sciences and
2Department of Community & Health Sciences, Aga Khan University, Karachi, Pakistan.
3Department of Biomedical Engineering, University of Virginia, Charlottesville, Virginia, USA.
4Department of Pediatrics & Child Health, Aga Khan University, Karachi, Pakistan.
5Division of Infectious Diseases and International Health, Department of Medicine, University of Virginia School of Medicine, Charlottesville, Virginia, USA.
6Department of Public Health Sciences, University of Virginia, Charlottesville, Virginia, USA.
7Division of Gastroenterology, Hepatology and Nutrition, Cincinnati Children’s Hospital, Cincinnati, Ohio, USA.
Address correspondence to: Syed Asad Ali, Chair, Department of Community Health Sciences, Professor, Pediatric Infectious Disease, Aga Khan University, National Stadium Road PO Box 3500, Karachi 74800, Pakistan. Email: asad.ali@aku.edu. Or to: Sean Ryan Moore, Division Director, Division of Gastroenterology, Hepatology and Nutrition, Professor, UC Department of Pediatrics, Cincinnati Children’s Hospital Medical Center, 3333 Burnet Ave, Cincinnati, Ohio 45229, USA. Email: sean.moore@cchmc.org.
Authorship note: ZJ and GFH contributed equally to this work.
Find articles by Ahmed, K. in: PubMed | Google Scholar
1Department of Biological & Biomedical Sciences and
2Department of Community & Health Sciences, Aga Khan University, Karachi, Pakistan.
3Department of Biomedical Engineering, University of Virginia, Charlottesville, Virginia, USA.
4Department of Pediatrics & Child Health, Aga Khan University, Karachi, Pakistan.
5Division of Infectious Diseases and International Health, Department of Medicine, University of Virginia School of Medicine, Charlottesville, Virginia, USA.
6Department of Public Health Sciences, University of Virginia, Charlottesville, Virginia, USA.
7Division of Gastroenterology, Hepatology and Nutrition, Cincinnati Children’s Hospital, Cincinnati, Ohio, USA.
Address correspondence to: Syed Asad Ali, Chair, Department of Community Health Sciences, Professor, Pediatric Infectious Disease, Aga Khan University, National Stadium Road PO Box 3500, Karachi 74800, Pakistan. Email: asad.ali@aku.edu. Or to: Sean Ryan Moore, Division Director, Division of Gastroenterology, Hepatology and Nutrition, Professor, UC Department of Pediatrics, Cincinnati Children’s Hospital Medical Center, 3333 Burnet Ave, Cincinnati, Ohio 45229, USA. Email: sean.moore@cchmc.org.
Authorship note: ZJ and GFH contributed equally to this work.
Find articles by Mallawaarachchi, I. in: PubMed | Google Scholar
1Department of Biological & Biomedical Sciences and
2Department of Community & Health Sciences, Aga Khan University, Karachi, Pakistan.
3Department of Biomedical Engineering, University of Virginia, Charlottesville, Virginia, USA.
4Department of Pediatrics & Child Health, Aga Khan University, Karachi, Pakistan.
5Division of Infectious Diseases and International Health, Department of Medicine, University of Virginia School of Medicine, Charlottesville, Virginia, USA.
6Department of Public Health Sciences, University of Virginia, Charlottesville, Virginia, USA.
7Division of Gastroenterology, Hepatology and Nutrition, Cincinnati Children’s Hospital, Cincinnati, Ohio, USA.
Address correspondence to: Syed Asad Ali, Chair, Department of Community Health Sciences, Professor, Pediatric Infectious Disease, Aga Khan University, National Stadium Road PO Box 3500, Karachi 74800, Pakistan. Email: asad.ali@aku.edu. Or to: Sean Ryan Moore, Division Director, Division of Gastroenterology, Hepatology and Nutrition, Professor, UC Department of Pediatrics, Cincinnati Children’s Hospital Medical Center, 3333 Burnet Ave, Cincinnati, Ohio 45229, USA. Email: sean.moore@cchmc.org.
Authorship note: ZJ and GFH contributed equally to this work.
Find articles by
Ma, J.
in:
PubMed
|
Google Scholar
|
1Department of Biological & Biomedical Sciences and
2Department of Community & Health Sciences, Aga Khan University, Karachi, Pakistan.
3Department of Biomedical Engineering, University of Virginia, Charlottesville, Virginia, USA.
4Department of Pediatrics & Child Health, Aga Khan University, Karachi, Pakistan.
5Division of Infectious Diseases and International Health, Department of Medicine, University of Virginia School of Medicine, Charlottesville, Virginia, USA.
6Department of Public Health Sciences, University of Virginia, Charlottesville, Virginia, USA.
7Division of Gastroenterology, Hepatology and Nutrition, Cincinnati Children’s Hospital, Cincinnati, Ohio, USA.
Address correspondence to: Syed Asad Ali, Chair, Department of Community Health Sciences, Professor, Pediatric Infectious Disease, Aga Khan University, National Stadium Road PO Box 3500, Karachi 74800, Pakistan. Email: asad.ali@aku.edu. Or to: Sean Ryan Moore, Division Director, Division of Gastroenterology, Hepatology and Nutrition, Professor, UC Department of Pediatrics, Cincinnati Children’s Hospital Medical Center, 3333 Burnet Ave, Cincinnati, Ohio 45229, USA. Email: sean.moore@cchmc.org.
Authorship note: ZJ and GFH contributed equally to this work.
Find articles by Aziz, F. in: PubMed | Google Scholar
1Department of Biological & Biomedical Sciences and
2Department of Community & Health Sciences, Aga Khan University, Karachi, Pakistan.
3Department of Biomedical Engineering, University of Virginia, Charlottesville, Virginia, USA.
4Department of Pediatrics & Child Health, Aga Khan University, Karachi, Pakistan.
5Division of Infectious Diseases and International Health, Department of Medicine, University of Virginia School of Medicine, Charlottesville, Virginia, USA.
6Department of Public Health Sciences, University of Virginia, Charlottesville, Virginia, USA.
7Division of Gastroenterology, Hepatology and Nutrition, Cincinnati Children’s Hospital, Cincinnati, Ohio, USA.
Address correspondence to: Syed Asad Ali, Chair, Department of Community Health Sciences, Professor, Pediatric Infectious Disease, Aga Khan University, National Stadium Road PO Box 3500, Karachi 74800, Pakistan. Email: asad.ali@aku.edu. Or to: Sean Ryan Moore, Division Director, Division of Gastroenterology, Hepatology and Nutrition, Professor, UC Department of Pediatrics, Cincinnati Children’s Hospital Medical Center, 3333 Burnet Ave, Cincinnati, Ohio 45229, USA. Email: sean.moore@cchmc.org.
Authorship note: ZJ and GFH contributed equally to this work.
Find articles by Ali, S. in: PubMed | Google Scholar
1Department of Biological & Biomedical Sciences and
2Department of Community & Health Sciences, Aga Khan University, Karachi, Pakistan.
3Department of Biomedical Engineering, University of Virginia, Charlottesville, Virginia, USA.
4Department of Pediatrics & Child Health, Aga Khan University, Karachi, Pakistan.
5Division of Infectious Diseases and International Health, Department of Medicine, University of Virginia School of Medicine, Charlottesville, Virginia, USA.
6Department of Public Health Sciences, University of Virginia, Charlottesville, Virginia, USA.
7Division of Gastroenterology, Hepatology and Nutrition, Cincinnati Children’s Hospital, Cincinnati, Ohio, USA.
Address correspondence to: Syed Asad Ali, Chair, Department of Community Health Sciences, Professor, Pediatric Infectious Disease, Aga Khan University, National Stadium Road PO Box 3500, Karachi 74800, Pakistan. Email: asad.ali@aku.edu. Or to: Sean Ryan Moore, Division Director, Division of Gastroenterology, Hepatology and Nutrition, Professor, UC Department of Pediatrics, Cincinnati Children’s Hospital Medical Center, 3333 Burnet Ave, Cincinnati, Ohio 45229, USA. Email: sean.moore@cchmc.org.
Authorship note: ZJ and GFH contributed equally to this work.
Find articles by Moore, S. in: PubMed | Google Scholar
Published July 22, 2025 - More info
JCI Insight. 2025;10(14):e188993. https://doi.org/10.1172/jci.insight.188993.
© 2025 Jamil et al. This work is licensed under the Creative Commons Attribution 4.0 International License. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
Received: November 22, 2024; Accepted: June 3, 2025

-
Results
Participant characteristics.
This study includes participants selected from the parent study, SEEM, in Pakistan in which 416 infants were enrolled as cases (n = 365) and controls (n = 51) from a rural setting. Infants (0–1 month old) were screened by monitoring their weight and length monthly up to 6 months and were then classified as either well-nourished controls (WLZ ≥ 0, length-for-age z [LAZ] ≥-1, n = 51) or undernourished controls (wasted with WLZ ≤ –2, n = 365) based on at least 2 consecutive months of anthropometry. Of the 365 controls, wasted infants (WLZ < –2, n = 187) whose growth did not improve after their primary caregiver participated in an intensive nutritional education intervention were provided Acha Mum at the age of 9 months, a locally produced chickpea-based RUSF, daily for 8 weeks. The blood, fecal, and urine samples were collected prior to the initiation of RUSF, to determine EED biomarkers. These baseline characteristics and biomarkers are summarized in Table 1. Maternal and paternal BMI, as well as infant anthropometrics (0–1 month of age), were significantly lower in controls (P < 0.05), with differences that became even more pronounced by 9 months of age (P < 0.05). Breastfeeding patterns were similar between groups during the first 6 months of life. Of these 187 controls, 112 responded (41%), while 75 (59%) were classified as nonresponders as assessed by WLZ 1-week after intervention (10). To select 60 infants for this substudy (of 187 controls in SEEM), we calculated weight-for-age (WAZ) velocity (ΔWAZ/month) during the 8-week intervention rather than WLZ velocity, given that changes in infant length are more challenging to reliably measure and detect over an 8-week nutritional intervention relative to changes in infant weight.
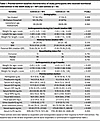 Table 1
Table 1Preintervention baseline characteristics of study participants who received nutritional intervention in the SEEM study (n = 187) and controls (n = 51)
Based on ΔWAZ, 30 best responders with ΔWAZ > 0.5/month and 30 worst responses with ΔWAZ ≤ 0/month were selected in addition to 28 controls (of the 51 controls from the parent study), as they lived in the same settings with similar socioeconomic statuses (Figure 1A). We performed fecal 16S sequencing of samples collected at 4 time points: (preintervention at 9 months, 1 week after intervention at 12 months, 1 month after intervention [13 months], and 6 months after intervention [18 months]) as shown in Figure 1B. Relabeling the controls based on their response, the baseline characteristics of the 88 participants in this sub-study are summarized in Supplemental Tables 1 and 2 (supplemental material available online with this article; https://doi.org/10.1172/jci.insight.188993DS1). These selected 88 participants were a true representation of the larger SEEM cohort (Supplemental Table 3).
 Figure 1
Figure 1Overview of study design. (A) Flow diagram of study population and protocols. (B) Infogram of study timelines and sample collection.
Ponderal growth responses of selected participants to RUSF administration.
Regarding anthropometrics, 30 best responders (RUSF-R) demonstrated a mean change in WAZ of 0.413 over the course of the intervention (Figure 2A and Table 2). RUSF-R showed significant improvements in overall nutritional status; SAM declined from 33% to 0%, and MAM declined from 63% to 20% (Figure 2B). In contrast, the 30 children with the highest decline in WAZ during the course of intervention (RUSF-NR) demonstrated either no improvements in WAZ or a decline, with a mean monthly WAZ velocity over the intervention of –0.081. The proportion of children with SAM increased in the RUSF-NR from 27% at baseline to 57% postintervention. WAZ and WLZ scores were slightly lower in RUSF-R at baseline, indicating RUSF response was neither due to better nutritional status at baseline nor regression to the mean. Of note, RUSF-R were less wasted on their first anthropometrics (0 to 1 month) than RUSF-NR (Supplemental Table 1). All other baseline indices were similar between groups, including RUSF adherence and illness burden (Supplemental Figure 1A).
 Figure 2
Figure 2Children selected for gut microbiome analysis show diverging yet durable responses to RUSF. (A) Mean change in WAZ score per month during RUSF. (B) Nutritional status of responders (n = 30) and nonresponders (n = 30) with 16S data before and after RUSF. (C) On average, responders had worse ponderal growth prior to RUSF but showed persistent growth improvements in response to RUSF as measured by weight-for-age z scores, length-for-age z scores, and weight-for-length z scores. Age- and region-matched well-nourished control cohort shown in blue (n = 28).
We tracked growth trajectories in RUSF-R and RUSF-NR beyond the completion of supplementation (Figure 2C). Improvements in WAZ and WLZ continued in RUSF-R after the intervention was completed, persisting until the end of follow-up at 24 months, indicating a durable RUSF effect. RUSF-NR did not respond immediately to nutritional intervention; however, we noted improvements in ponderal growth beginning at age 13 months (Table 2). Length-for-age z (LAZ) scores remained largely unchanged in both RUSF-R and RUSF-NR over the course of intervention and during follow-up. In controls who did not require nutritional intervention, anthropometric indices gradually declined during follow-up. Taken together, this suggests that RUSF was ultimately beneficial even for RUSF-NR.
Serum, urine, and fecal biomarkers distinguish wasted children from controls.
We hypothesized baseline growth, inflammation, and EED biomarkers would be distinct in wasted infants compared with well-nourished controls. These biomarker data available from the entire SEEM cohort (n = 235) were compared using Principal Component Analysis (PCA) to identify global group differences (Figure 3A). Our model was able to separate controls and controls along Principal Component 2 (PC2), with variation driven by biomarkers of growth and nutritional status (IGF1, prealbumin, GLP2, and leptin) as well as acute phase proteins C-reactive protein (CRP) and Alpha-1–acid glycoprotein (AGP) (Supplemental Figure 2A). Control samples clustered tightly, whereas controls displayed a more heterogeneous distribution along PC1, with a subset of participants designated as controls regardless of nutritional status. PC1 represented most of the variation within the model, primarily driven by inflammatory cytokines such as IL-1β, IFN-γ, and TNF-α.
 Figure 3
Figure 3Malnourished children exhibit increased systemic inflammation compared with healthy age-matched controls. (A) PCA of biomarker profiles from malnourished children (n = 148) and healthy aged-matched children (n = 39). (B–D) Urine (B), fecal (C), and serum biomarker/cytokine (D) levels from children at 9 months of age prior to RUSF in the SEEM cohort. *P < 0.05. Wilcoxin’s rank-sum test with Benjamini Hochburg corrections.
The PCA of the 88 study participants selected for this RUSF subanalysis (30 RUSF-R, 30 RUSF-NR, and 28 controls) showed a similar pattern of clustering (Supplemental Figure 2B), indicating that biomarker profiles from these subgroups were broadly representative of the total cohort.
PCA findings were confirmed by univariate analyses. Growth and nutritional status biomarkers, including IGF1, prealbumin, GLP2, and leptin, were significantly elevated in control samples compared with controls (Figure 3, B–D). Controls displayed evidence of systemic inflammation, with significantly higher concentrations of inflammatory biomarkers such as CRP, IL-6, IFN-γ, TNF-α, and IL-1β. Notably, the antiinflammatory cytokine IL-10 was higher in controls than in controls, suggesting feedback regulation. In addition, the urinary excretion of the intestinal tight junction protein claudin-15 was significantly higher in controls, consistent with previous associations of urinary claudin-15 with decreased gut barrier function observed in EED (11, 12). Fecal markers of increased intestinal inflammation were similar across controls and controls: fecal MPO and NEO levels were similar between groups, and fecal lipocalin-2 (LCN2) was elevated in controls compared with controls.
Preintervention biomarkers predict RUSF response.
The heterogeneity of biomarker profiles observed in controls suggests a mixed population among this group. Therefore, we hypothesized that biomarkers would help identify which undernourished infants were less likely to respond to RUSF. Relabeling controls by nutritional response (RUSF-R versus RUSF-NR) supported this hypothesis; most samples that failed to cluster with control samples belonged to the nonresponder group (Figure 4A). Heterogeneous samples were more likely to be RUSF-NR; however, RUSF-R and RUSF-NR groups did not cleanly separate, indicating overall broad similarity in biomarker profiles. In univariate analyses, fecal NEO, serum CRP, and IL-10 were higher in RUSF-NR versus RUSF-R before intervention. However, after multiple testing corrections, there were no significant differences observed between controls and controls (Figure 4, B–D). Therefore, we took a machine learning approach to identify biomarkers that are more predictive of RUSF response.
 Figure 4
Figure 4Baseline inflammatory biomarkers predict response to intervention. (A) PCA displaying the relationship between biomarkers and cytokines in the responders (n = 66), nonresponders (n = 82), and controls (n = 38) at 9 months of age. (B–D) Urine (B), fecal (C), and serum biomarker (D) levels from stunted children at 9 months of age prior to RUSF in SEEM cohort. *P < 0.05. (E) Variable importance as shown by mean average decrease in GINI for each biomarker in a random forest model trained to predict whether a child would respond to the nutritional intervention. Random forest had a 73% CV accuracy. Circles indicate average importance; data are shown as mean ± SD of 20-fold cross validation. (F) Accuracy of logistic regression models trained to predict response from individual biomarkers or combinations of biomarkers deemed important from random forest analysis.
Biomarkers were incorporated into an ensemble of random forest (RF) models with 20-fold cross-validation. Using this approach, biomarker data were able to correctly predict response to nutritional intervention with 60% accuracy (Figure 4E). A distinct set of 9 features (GLP2, MPO, prealbumin, NEO, claudin-15, creatinine, CRP, IL-10, and MCP1) representing a mix of growth and inflammatory biomarkers was identified as most important for distinguishing responders from nonresponders. We observed a trend toward increased markers of growth and nutritional status (GLP2, prealbumin, creatinine) in responders, consistent with the elevation of these markers in control samples relative to controls. We also noted a trend toward increased intestinal inflammation (MPO, NEO) in nonresponders — a difference not seen in comparing controls and controls. IL-10 was significantly higher in responders and CRP was significantly higher in nonresponders, highlighting elevated systemic inflammation as predictive of failure to respond. A logistic regression model was trained based on these 9 biomarkers to interrogate the importance of individual biomarkers. This model correctly predicted response with 68% accuracy on training set data and 73% accuracy on withheld test set data (Figure 4F). Regression models were built using either individual or the collective top 4 biomarkers according to RF performed poorly, with accuracy ranging from 50% to 60%. Broadly, these results indicate that biomarkers of growth, in conjunction with biomarkers of intestinal and systemic inflammation, can reliably distinguish RUSF responders and nonresponders prior to intervention.
Gut microbiome diversity differs between wasted children and controls.
Next, we explored the fecal microbiome community profiles of controls (including responders and nonresponders) and controls. We performed 16S rRNA sequencing on stool samples collected at baseline (preintervention) and at several time points after intervention to identify amplicon sequence variants (ASVs), representing unique biological sequences that can be assigned a taxonomy. α Diversity was significantly higher in controls as compared with controls (Figure 5A). This was true when measuring observed ASVs as a measure of richness (how many unique taxa are present) and when measuring Simpson’s diversity, which accounts for both richness and evenness (how evenly the relative abundance of taxa are distributed). Controls and controls also displayed significantly different β diversity, as visualized using nonparametric multidimensional scaling (NMDS) on a Bray-Curtis dissimilarity matrix (Figure 5B). These results suggest that wasted children in our cohort had a distinct microbiome from controls, characterized by increased diversity and greater evenness of taxa.
 Figure 5
Figure 5Wasted microbiota are more diverse relative to well-nourished microbiota. (A) α Diversity measurements (number of observed species, Simpson index) of wasted controls and healthy controls at 9 months of age prior to RUSF. (B) Nonmetric multidimensional scaling plot on Bray-Curtis matrices comparing the community composition of the controls and controls prior to the intervention. (C and D) Relative abundance of phylum (C) and class (D) level taxonomy between controls and controls prior to the nutritional intervention. *P < 0.05. An OPLS-DA model was constructed to discriminate between controls and controls using the relative abundance of ASVS from their fecal microbiome. The model outperformed all of 1,000 randomly permuted models (P < 0.001). (E) Scatter plot of the X scores on latent variables 1 and 2 (LV1 and LV2), where each point represents 1 sample. (F) Bar plot shows the variable importance in projection (VIP) scores, artificially oriented in the direction of loadings on LV1, and colored according to their association with patient or control samples, labeled with genus name and ASV number. VIP scores > 1 indicate a variable with greater than average influence on the projection.
We next determined which specific phylogenetic groups were different between controls and controls. Consistent with the decreased diversity observed in controls versus controls, the community composition of control samples was largely dominated by Actinomycetota, whereas this phylum made up a significantly smaller proportion of the total community in controls (Figure 5C). The lower relative abundance of Actinomycetota in controls was compensated by a significant increase in Pseudomonadota and trends toward increased Bacillota and Bacteroidota relative to controls. These changes were also reflected at the class level (Figure 5D), with a relative decrease in Actinomycetia in controls and increased Gammaproteobacteria and trends toward increased Bacteroidia, Negativicutes, and Clostridia.
Finally, a LASSO-regularized, orthogonal partial least squares-discriminant analysis (OPLS-DA) model was constructed to identify ASVs that best discriminate between groups (Figure 5E). The model outperformed all 1,000 randomly permuted models (P < 0.001) and achieved a cross-validation accuracy of 78% (Supplemental Figure 3A). Variable importance in projection (VIP) scores were plotted for top features and reflected ASV important for discriminating between groups (absolute value of VIP > 1 is an important feature, with the sign of VIP indicating which group it is related to) (Figure 5F). This analysis identified ASV1620, a member of the Bifidobacterium genus, as particularly important in discriminating between controls and controls. Univariate analysis showed ASV1620 was significantly enriched in control samples (Supplemental Figure 3B). This ASV is a member of the Actinomycetota phylum and Bifidobacterium genus, which are known to be important in early life (9). As LASSO hides features that are linearly correlated to key driver features, ASVs with a high correlation to those identified in the model may also be significant and are shown in a correlation network (Supplemental Figure 3C). Taken together, our results show greater microbiome diversity at the expense of Bifidobacteria species is a signature feature of undernourished controls prior to RUSF.
ASV-level differences reliably distinguish responders from nonresponders among wasted children.
We next sought to identify compositional differences in gut microbial communities of RUSF responders and nonresponders — both baseline differences and longitudinal changes after intervention. We observed no significant differences in either α or β diversity between responder and nonresponders prior to RUSF (Figure 6, A and B), indicating the gut microbiota of these groups are broadly more similar to one another, relative to more striking diversity differences between controls and controls. Community composition in RUSF-NR mostly consisted of enrichment in Actinomycetota, whereas RUSF-R exhibited a significantly lower proportion of Actinomycetota and increased Pseudomonadota (Figure 6, C and D), similar to the differences we observed in controls versus controls. Parallel trends were seen at the class level, where the relative abundance of Actinomycetia and Gammaproteobacteria were significantly higher and lower in RUSF-R, respectively. In addition, we noted trends toward a higher relative baseline abundance of Bacillota at the phylum level and Negativicutes at the class level.
 Figure 6
Figure 6Responders and nonresponders harbor distinct fecal microbiota prior to RUSF. (A) α Diversity measurements (number of observed species, Simpson index) of responders and nonresponders at 9 months of age prior to RUSF. (B) Nonmetric multidimensional scaling plot on Bray-Curtis matrices comparing the community composition of the responders and nonresponders prior to the intervention. (C and D) Relative abundance of phylum (C) and Class level (D) taxonomy between controls and controls prior to the nutritional intervention. An OPSL-DA model was constructed to discriminate between responders and nonresponders using the relative abundance of ASVS from their fecal microbiome. The model outperformed all of 1,000 randomly permuted models (P < 0.001). (E) Scatter plot of the X scores on latent variables 1 and 2 (LV1 and LV2), where each point represents one sample. (F) Bar plot shows the variable importance in projection (VIP) scores, artificially oriented in the direction of loadings on LV1 and colored according to their association with responder or nonresponder samples, labeled with genus name and ASV number. VIP scores > 1 indicate a variable with greater than average influence on the projection.
We then generated a LASSO-regularized, OPLS-DA model to distinguish responders from nonresponders based on gut microbiome composition prior to RUSF. Remarkably, this model outperformed all 1,000 randomly permuted models (P < 0.001) and achieved a cross-validation accuracy of 93%, indicating distinct differences at the ASV level between RUSF-R and RUSF-NR (Supplemental Figure 4A). VIP scores were plotted and used to determine ASVs important for discriminating between groups (Figure 6, E and F), with univariate analysis showing significant enrichment in several of the important features (Supplemental Figure 4B). ASVs with a high correlation (70%) to those identified in the model that may also be of significance are shown in a heatmap (Supplemental Figure 4C). This analysis points to a distinct pre-RUSF microbiome state characterized by increased colonization of Pseudomonadota in children whose ponderal growth markedly improved after RUSF.
Responders display shifts in gut microbiome composition during RUSF, and microbiome composition in responders and nonresponders converge in the aftermath of RUSF.
We next turned our attention to longitudinal changes in the microbiome over RUSF supplementation. α Diversity increased during the intervention among responders and nonresponders (Figure 7A), a pattern also observed in controls (Supplemental Figure 5A). We identified no significant shifts in RUSF-NR microbiota at the phylum or class levels. In contrast, in RUSF-R, we identified a significant decline in the relative abundance of Pseudomonadota, largely driven by reductions in Gammaproteobacteria and an increase in Clostridia. This decline in Gammaproteobacteria may explain the ability of responders to gain weight during the intervention as pathogens such as E. coli, Shigella, and Salmonella are implicated in EED, contributing to malnutrition while disrupting intestinal function. Importantly, we observed a robust increase in the relative abundance of Negativicutes in RUSF-R relative to RUSF-NR (Supplemental Figure 5B) before and after RUSF (Figure 7B and Supplemental Figure 7C). In RUSF-NR, taxa were not immediately different before versus after intervention. At the family level, shifts in the Negativicutes class were driven by members of Veillonellaceae, Pseudomonadota by members of Enterobacteriaceae, and Actinomycetia by members of the Bifidobacteriaceae family (Figure 7C).
 Figure 7
Figure 7RUSF responders exhibit distinct shifts in gut microbiome composition over the course of the nutritional intervention. (A) α Diversity measurements (number of observed species, Simpson index) of responders and nonresponders before and after intervention. (B) Comparison of relative abundance of microbial taxa at the phylum level in responder and nonresponder stool samples over the course of the study. (C) The –log10 adjusted P values from a mixed linear effects model comparing the differences in the relative abundance at the class level between responders and nonresponders. The dotted line indicates an adjusted P < 0.05. (D) Scatter plot of the X scores on latent variables 1 and 2 (LV1 and LV2) from an OPSL-DA model trained to discern differences in ASV relative abundance between responders and nonresponders after intervention, where each point represents 1 sample. (E) Bar plot shows the variable importance in projection (VIP) scores, artificially oriented in the direction of loadings on LV1 and colored according to their association with responder or nonresponder samples. VIP scores > 1 indicate a variable with greater than average influence on the projection.
To identify specific ASVs associated with RUSF response, we constructed a LASSO regularized OPLS-DA model to discriminate between RUSF-R and RUSF-NR samples immediately after intervention (12 months of age) (Figure 7D). The model outperformed all of 1,000 randomly permuted models (P < 0.001) and achieved a cross-validation accuracy of 98% (Supplemental Figure 6A). A plot of VIP scores showed several ASVs as important in discriminating between responder and nonresponder samples after intervention. ASV2304, a member of the Lachnoclostridium genus, was the most important feature of RUSF-NR samples (Figure 7E). Univariate analysis showed this ASV to be present significantly more often in nonresponder samples than in responder samples. The most important microbial feature relevant to RUSF-R samples was ASV578, a member of the Veillonella genus and Negativicutes class that was present more abundantly in responders (Supplemental Figure 6B). ASVs with a high correlation (>70%) to those identified in the model that may also be of significance are shown in a heatmap (Supplemental Figure 6C). Together, this analysis shows a RUSF-driven transformation of the gut microbiome in RUSF-R distinct from that observed in RUSF-NR over the course of the nutritional intervention. This transformation is characterized by a robust decrease in Gammaproteobacteria and an increase in Negativicutes.
Systemic inflammation identifies RUSF-NR outliers with responder-like microbiome profiles.
Among the 30 worst RUSF-NR, we identified 7 individual nonresponders with distinct biomarker profiles relative to either controls or RUSF-R. These profiles were primarily driven by a significant elevation in biomarkers of systemic inflammation (Figure 8A). We next asked whether these individuals harbored distinct microbiota relative to other RUSF-NR. The microbiomes of inflamed RUSF-NR showed greater similarity to RUSF-R than to noninflamed RUSF-NR (Figure 8B). This was most evidenced by lower levels of Actinomycetota and increased levels of Pseudomonadota in inflamed versus noninflamed RUSF-NR. Inflamed RUSF-NR also exhibited distinct growth trajectories, with higher WAZ scores prior to RUSF followed by marked decreases in WAZ during the intervention (Figure 8C). Follow-up analysis also showed that these infants experienced higher rates of diarrheal episodes and acute respiratory infections (Supplemental Figure 7, A and B, Supplemental Table 3). These findings suggest that microbiome structure and function play an important role in RUSF response; however, systemic inflammation and acute illnesses can undermine that response.
 Figure 8
Figure 8RUSF nonresponders with high levels of inflammation harbor fecal microbiota that resemble those of responders. (A) PCA displaying the relationship between biomarkers and cytokines measured before intervention from patients with matching microbiome data in the responders (n = 30), nonresponders (n = 30), and control groups (n = 28). Black circles denote nonresponder samples with high levels of inflammation selected for downstream analysis. (B) Comparison of the average relative abundance of microbial taxa at the phylum level in responder, nonresponder, and inflamed nonresponder stool samples prior to the nutritional intervention. (C) Longitudinal change in WAZ score for control, responder, nonresponder, and inflamed nonresponder children over the first 2 years of life.
Validation of the predictive model.
After pinpointing microbial taxa with strong predictive potential, we validated our findings in an independent Bangladeshi cohort of young children. Since limited studies were suitable for validation, we selected the 2022 study by Chen et al. (13) due to its utilization of a chickpea-based MDCF. We utilized the data of 60 children who received the MDCF and categorized them into 3 groups based on changes in WAZ during the intervention (Supplemental Figure 8A). The fecal microbiome was compared between children showing the most favorable response and those showing the least. Unlike our dataset, at the preintervention stage, significant differences in Pseudomonadota or Negativicutes abundance in the Chen et al.(13) study were not seen (Supplemental Figure 8B). To uncover any consistent microbial taxa with predictive potential across cohorts, we first used PCA on preintervention composition data (Supplemental Figure 8C) and, seeing no significant batch effects, trained an Orthogonal Partial Least Squares Discriminant Analysis (OPLS-DA) model using preintervention microbiome data from the SEEM study and applied it to the preintervention validation dataset (Supplemental Figure 8D). Although our model identified several taxa significantly linked to response, it only classified patients with 54% accuracy in the validation dataset. While this accuracy is only marginally better than chance, it underscores the microbiome composition variations between cohorts and studies.












