Clinical Research and Public HealthMetabolism
Open Access | ![]() 10.1172/jci.insight.146000
10.1172/jci.insight.146000
Metabolic flexibility across the spectrum of glycemic regulation in youth
Fida Bacha,1,2 Sara Klinepeter Bartz,1,2 Maurice Puyau,1 Anne Adolph,1 and Susan Sharma1
1United States Department of Agriculture Agricultural Research Service (USDA/ARS) Children’s Nutrition Research Center, Texas Children’s Hospital, Baylor College of Medicine, Houston, Texas, USA.
2Division of Pediatric Endocrinology and Diabetes, Texas Children’s Hospital, Baylor College of Medicine, Houston, Texas, USA.
Address correspondence to: Fida Bacha, USDA/ARS Children’s Nutrition Research Center, Baylor College of Medicine, 1100 Bates Street, Houston, Texas 77030, USA. Phone: 713.798.7166; Email: fbacha@bcm.edu.
Find articles by
Bacha, F.
in:
PubMed
|
Google Scholar
|

1United States Department of Agriculture Agricultural Research Service (USDA/ARS) Children’s Nutrition Research Center, Texas Children’s Hospital, Baylor College of Medicine, Houston, Texas, USA.
2Division of Pediatric Endocrinology and Diabetes, Texas Children’s Hospital, Baylor College of Medicine, Houston, Texas, USA.
Address correspondence to: Fida Bacha, USDA/ARS Children’s Nutrition Research Center, Baylor College of Medicine, 1100 Bates Street, Houston, Texas 77030, USA. Phone: 713.798.7166; Email: fbacha@bcm.edu.
Find articles by Bartz, S. in: PubMed | Google Scholar
1United States Department of Agriculture Agricultural Research Service (USDA/ARS) Children’s Nutrition Research Center, Texas Children’s Hospital, Baylor College of Medicine, Houston, Texas, USA.
2Division of Pediatric Endocrinology and Diabetes, Texas Children’s Hospital, Baylor College of Medicine, Houston, Texas, USA.
Address correspondence to: Fida Bacha, USDA/ARS Children’s Nutrition Research Center, Baylor College of Medicine, 1100 Bates Street, Houston, Texas 77030, USA. Phone: 713.798.7166; Email: fbacha@bcm.edu.
Find articles by Puyau, M. in: PubMed | Google Scholar
1United States Department of Agriculture Agricultural Research Service (USDA/ARS) Children’s Nutrition Research Center, Texas Children’s Hospital, Baylor College of Medicine, Houston, Texas, USA.
2Division of Pediatric Endocrinology and Diabetes, Texas Children’s Hospital, Baylor College of Medicine, Houston, Texas, USA.
Address correspondence to: Fida Bacha, USDA/ARS Children’s Nutrition Research Center, Baylor College of Medicine, 1100 Bates Street, Houston, Texas 77030, USA. Phone: 713.798.7166; Email: fbacha@bcm.edu.
Find articles by Adolph, A. in: PubMed | Google Scholar
1United States Department of Agriculture Agricultural Research Service (USDA/ARS) Children’s Nutrition Research Center, Texas Children’s Hospital, Baylor College of Medicine, Houston, Texas, USA.
2Division of Pediatric Endocrinology and Diabetes, Texas Children’s Hospital, Baylor College of Medicine, Houston, Texas, USA.
Address correspondence to: Fida Bacha, USDA/ARS Children’s Nutrition Research Center, Baylor College of Medicine, 1100 Bates Street, Houston, Texas 77030, USA. Phone: 713.798.7166; Email: fbacha@bcm.edu.
Find articles by Sharma, S. in: PubMed | Google Scholar
Published February 22, 2021 - More info
JCI Insight. 2021;6(4):e146000. https://doi.org/10.1172/jci.insight.146000.
© 2021 Bacha et al. This work is licensed under the Creative Commons Attribution 4.0 International License. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
Received: November 13, 2020; Accepted: January 13, 2021
-
Results
Physical and metabolic characteristics of study participants. The participants with NW and with obesity across the glycemic spectrum did not differ with respect to age, sex distribution, or Tanner stage. They were all pubertal, Tanner stages II–V (Table 1). There was a difference in race/ethnicity among the groups, with more Hispanic youth with type 2 diabetes. Therefore, we adjusted for race/ethnicity in subsequent analyses of metabolic variables. BMI z score, and measures of total body adiposity (%body fat, fat mass) or abdominal fat (waist circumference or visceral fat), were significantly higher in the youth with obesity compared with the NW group as expected, with no significant differences in these parameters among the NGT, prediabetes, or type 2 diabetes groups. HbA1c was significantly higher in the type 2 diabetes group compared with the NW and NGT groups. Adiponectin concentrations were significantly lower in the type 2 diabetes and prediabetes compared with NW and in the type 2 diabetes compared with the obese NGT group, after adjusting for race (given race-related differences in adiponectin) (20).
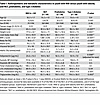 Table 1
Table 1Anthropometric and metabolic characteristics in youth with NW versus youth with obesity and NGT, prediabetes, and type 2 diabetes
Substrate utilization and metabolic flexibility across the glycemic spectrum. RER was not significantly different among the 4 groups at baseline. Postabsorptive fat oxidation was not significantly different among the 4 groups before and after adjustment for race, sex, and Tanner stage (Table 2). After further adjustment for fat mass and lean mass, fat oxidation was lower in the NW compared with the other 3 groups (0.66 ± 0.09 mg/kg/min in NW vs. 0.91 ± 0.07 mg/kg/min in NGT, 0.86 ± 0.06 mg/kg/min in the prediabetes, and 0.97 ± 0.06 mg/kg/min in the type 2 diabetes groups, respectively, P = 0.004). Postabsorptive glucose oxidation was higher in the NW group compared with the other 3 groups. This difference persisted after further adjustment for fat mass and lean mass (P = 0.04).
 Table 2
Table 2Indirect calorimetry and metabolic parameters in the fasting postabsorptive state and at the steady state of the hyperinsulinemic-euglycemic clamp in youth with NW and with obesity across the spectrum of glycemic regulation
Despite higher plasma fasting insulin concentrations, the rate of lipolysis was not significantly lower in the groups with obesity compared with the NW group; thus, adipose tissue insulin sensitivity was significantly lower in the groups with obesity across the spectrum of glycemia compared with the NW group (Table 2). Fasting FFA concentrations were significantly higher in the obese groups with dysglycemia compared with the other 2 groups (NW and obese NGT) (P < 0.001) (Table 2) and inversely related to adipose tissue insulin sensitivity (r = –0.46, P < 0.001).
The groups with dysglycemia had less increase in glucose oxidation and less suppression in fat oxidation in response to hyperinsulinemia compared with the NGT and NW groups (Table 2). Metabolic flexibility (ΔRER) decreased from NW to obese with NGT, prediabetes, and type 2 diabetes groups (0.10 ± 0.04, 0.08 ± 0.04, 0.07 ± 0.04, and 0.06 ± 0.04 respectively, P = 0.006). ΔRER remained significantly lower in the type 2 diabetes and prediabetes groups compared with NW and in the type 2 diabetes compared with the NGT group, after adjusting for race, sex, and Tanner stage. ΔRER was not significantly different between the NW and obese NGT groups (Figure 1).
 in youth with obesity across the spectrum of glycemic regulation [NGT (n = 24), prediabetes (n = 28), and type 2 diabetes (n = 28)] compared with NW (n = 24) peers.") Figure 1
Figure 1Metabolic flexibility (ΔRER) in youth with obesity across the spectrum of glycemic regulation [NGT (n = 24), prediabetes (n = 28), and type 2 diabetes (n = 28)] compared with NW (n = 24) peers. The box plots depict the minimum and maximum values (whiskers), the upper and lower quartiles, the median, and the mean (circle). The length of the box represents the interquartile range. One-way ANOVA. a, post hoc Bonferroni’s P < 0.05 in prediabetes vs. NW and in type 2 diabetes vs. NW; b, post hoc Bonferroni’s P < 0.05 in type 2 diabetes vs. NGT.
Under insulin-stimulated conditions, the oxidative and nonoxidative glucose disposal rates were significantly lower in the obese groups compared with NW and lower in the dysglycemia groups compared with obese NGT. Peripheral insulin sensitivity (total and adjusted per FFM) decreased across the spectrum of glycemia in the groups with obesity and was significantly lower than that of the NW group (Table 2). FFA concentrations continued to be significantly higher in the group with type 2 diabetes compared with the other 3 groups under the insulin-stimulated conditions of the HEC (Table 2).
Relationship of metabolic flexibility to abdominal adiposity, HbA1c, adiponectin, and lipids. ΔRER was not related to total body fat, fat mass, or lean mass but was inversely related to waist circumference (r = –0.23, P = 0.048) and to visceral adipose tissue (VAT) (r = –0.27, P = 0.02). The relationship to VAT was not significant (r = –0.21, P = 0.07) after adjusting for race. ΔRER was inversely related to HbA1c (r = –0.2, P = 0.049), triglycerides (r = –0.28, P = 0.04), and triglyceride/HDL-cholesterol ratio (r = –0.3, P = 0.002) and positively related to HDL-cholesterol (r = 0.2, P = 0.04) and adiponectin (r = 0.37, P < 0.001).
Relationship of metabolic flexibility to glucose and lipid metabolism. After adjusting for race, sex, and Tanner stage, fasting FFA concentration was inversely related to baseline and clamp steady-state glucose oxidation (r = –0.36, P < 0.001 for both). Fasting FFA was inversely related to fasting RER (r = –0.31, P = 0.002). Clamp FFA concentration was inversely related to insulin-stimulated glucose oxidation (r = –0.28, P = 0.005) and positively to fat oxidation (r = 0.21, P = 0.04).
After adjusting for race, sex, and Tanner stage, ΔRER was positively related to clamp steady-state glucose oxidation (r = 0.49, P < 0.001) and nonoxidative glucose disposal (r = 0.2, P = 0.06) and inversely related to fat oxidation (r = –0.44, P < 0.001). ΔRER also correlated positively with percentage suppression of FFA in response to hyperinsulinemia (r = 0.3, P = 0.01).
ΔRER correlated positively with insulin-stimulated glucose disposal (RdFFM) (r = 0.35, P < 0.001) and with insulin sensitivity per FFM before (r = 0.35, P < 0.001) and after (r = 0.29, P = 0.003) adjusting for race, sex, and Tanner stage (Figure 2). The relationship of ΔRER to adipose tissue insulin sensitivity (IS) was not significant (r = 0.2, P = 0.08).
 in youth with NW (n = 24) and with obesity across the spectrum of glycemic regulation [NGT (n = 24), prediabetes (n = 28), and type 2 diabetes (n = 28)].") Figure 2
Figure 2Relationship of insulin sensitivity to metabolic flexibility (ΔRER) in youth with NW (n = 24) and with obesity across the spectrum of glycemic regulation [NGT (n = 24), prediabetes (n = 28), and type 2 diabetes (n = 28)].
To identify significant independent determinants of metabolic flexibility, we performed multiple regression analysis. In a model with ΔRER as the dependent variable, ISFFM (β = 0.4, P = 0.004) (or RdFFM [β = 0.30, P = 0.004]), percentage suppression of FFA (r = 0.26, P = 0.007), and race/ethnicity (β = –0.23, P = 0.02) contributed to the variance in ΔRER (R2 = 0.30, P < 0.001) independent of percentage body fat (or VAT), sex, Tanner stage, and HbA1c as covariates in the model.


 in youth with obesity across the spectrum of glycemic regulation [NGT (n = 24), prediabetes (n = 28), and type 2 diabetes (n = 28)] compared with NW (n = 24) peers.")
 in youth with NW (n = 24) and with obesity across the spectrum of glycemic regulation [NGT (n = 24), prediabetes (n = 28), and type 2 diabetes (n = 28)].")


